
 Submit a Paper
Submit a Paper Propose a Special lssue
Propose a Special lssue Open Access
Open Access
ARTICLE

The superiority of PMFs on reversing drug resistance of colon cancer and the effect on aerobic glycolysis-ROS-autophagy signaling axis
1 Department of Pharmacy, Guangdong Provincial Key Laboratory of Major Obstetric Diseases, Guangdong Provincial Clinical Research Center for Obstetrics and Gynecology, The Third Affiliated Hospital of Guangzhou Medical University, Guangzhou, 510150, China
2 School of Pharmaceutical Sciences, Guangzhou Medical University, Guangzhou, 511436, China
3 Department of Pharmacy, The Fourth Affiliated Hospital of Guangzhou Medical University, Guangzhou, 511300, China
* Corresponding Authors: YONGHENG ZHOU. Email: ; SENLING FENG. Email:
# These authors contributed equally to this work and shared the first authorship
(This article belongs to the Special Issue: New Insights in Drug Resistance of Cancer Therapy: A New Wine in an Old Bottle)
Oncology Research 2024, 32(12), 1891-1902. https://doi.org/10.32604/or.2024.048778
Received 18 December 2023; Accepted 28 February 2024; Issue published 13 November 2024
 View Full Text
View Full Text Download PDF
Download PDFAbstract
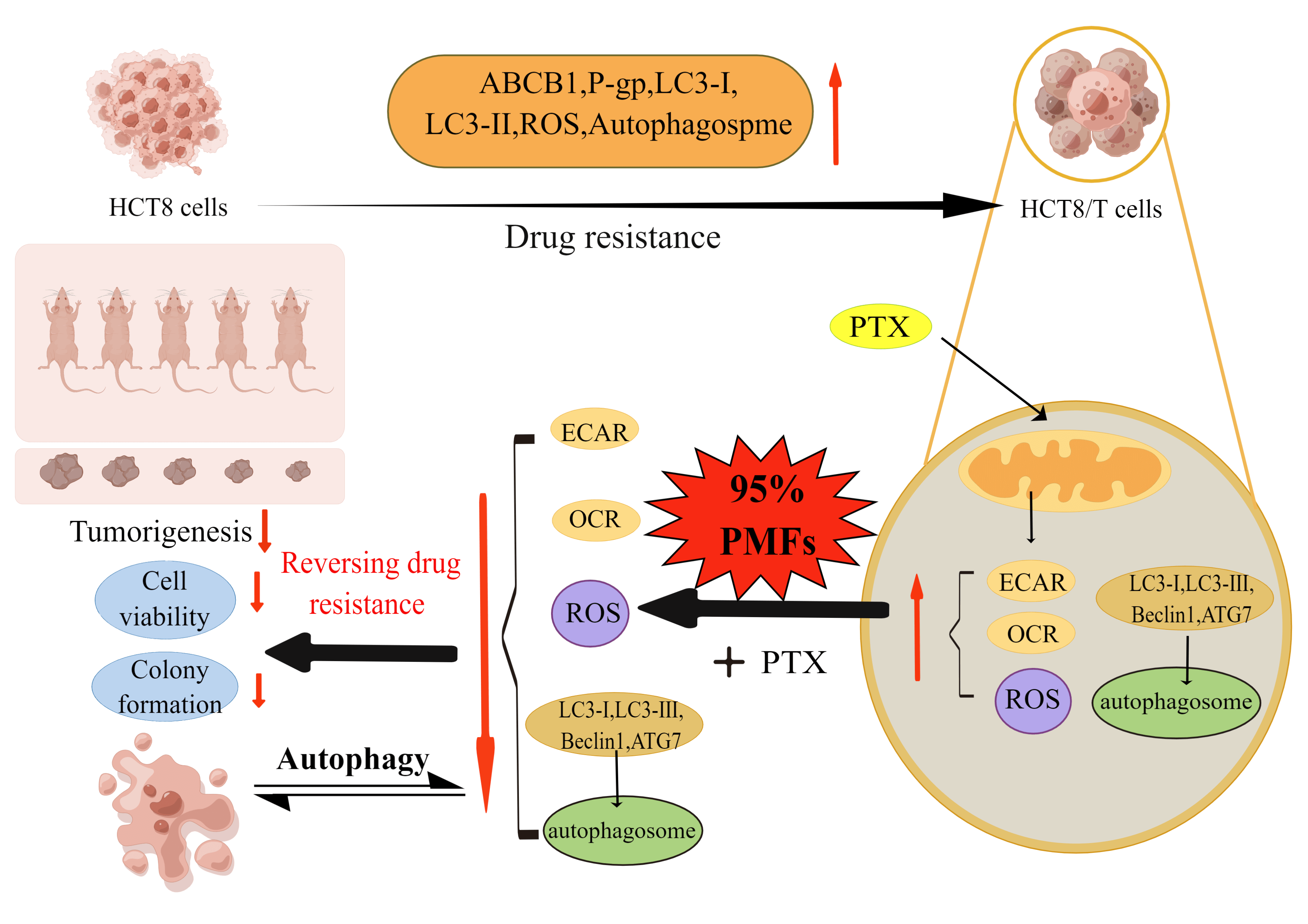
Background: Polymethoxylated flavones (PMFs) are compounds present in citrus peels and other Rutaceae plants, which exhibit diverse biological activities, including robust antitumor and antioxidant effects. However, the mechanism of PMFs in reversing drug resistance to colon cancer remains unknown. In the present study, we aimed to investigate the potential connection between the aerobic glycolysis-ROS-autophagy signaling axis and the reversal of PTX resistance in colon cancer by PMFs. Methods: MTT Cell viability assay and colony formation assay were used to investigate the effect of PMFs combined with PTX in reversing HCT8/T cell resistance ex vivo; the mRNA and protein levels of the target were detected by SDS-PAGE (sodium dodecyl sulfate-polyacrylamide gel electrophoresis), quantitative real-time fluorescence polymerase chain reaction (qRT-PCR) and Western blot protein immunoblotting (WB); An HCT8/T cell xenograft model was established to investigate the MDR reversal activity of PMFs in vivo; The extracellular acidification rate (ECAR) and the oxygen consumption rate (OCR) were detected to assess the cellular oxygen consumption rate and glycolytic process. Results: HCT8/T cells demonstrated significant resistance to PTX, up-regulating the expression levels of ABCB1 mRNA, P-gp, LC3-I, and LC3-II protein, and increasing intracellular reactive oxygen species (ROS) content. PMFs mainly contain two active ingredients, nobiletin, and tangeretin, which were able to reverse drug resistance in HCT8/T cells in a concentration-dependent manner. PMFs exhibited high tolerance in the HCT8/T nude mouse model while increasing the sensitivity of PTX-resistant cells and suppressing tumor growth significantly. PMFs combined with PTX reduced extracellular acidification rate (ECAR) and oxygen consumption rate (OCR) in HCT8/T cells. Additionally, PMFs reduced intracellular ROS content, down-regulated the expression levels of autophagy-related proteins LC3-I, LC3-II, Beclin1, and ATG7, and significantly reduced the number of autophagosomes in HCT8/T cells. Conclusions: The present study demonstrated that PMFs could potentially reverse PTX resistance in colon cancer by regulating the aerobic glycolysis-ROS-autophagy signaling axis, which indicated that PMFs would be potential potentiators for future chemotherapeutic agents in colon cancer.Graphic Abstract
Keywords
Cite This Article
 Copyright © 2024 The Author(s). Published by Tech Science Press.
Copyright © 2024 The Author(s). Published by Tech Science Press.This work is licensed under a Creative Commons Attribution 4.0 International License , which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
 Downloads
Downloads
 Citation Tools
Citation Tools