
 Submit a Paper
Submit a Paper Propose a Special lssue
Propose a Special lssue Open Access
Open Access
ARTICLE
Analysis of the role of dihydromyricetin derived from vine tea (Ampelopsis grossedentata) on multiple myeloma by activating STAT1/RIG-I axis
1 Department of Hematology, Shaoxing Shangyu People’s Hospital, Shaoxing, 312000, China
2 Department of Hematology, Zhuji People’s Hospital, Shaoxing, 311800, China
* Corresponding Author: MEI ZHOU. Email:
(This article belongs to the Special Issue: Signaling Pathway Crosstalk in Malignant Tumors: Molecular Targets and Combinatorial Therapeutics)
Oncology Research 2024, 32(8), 1359-1368. https://doi.org/10.32604/or.2024.043423
Received 02 July 2023; Accepted 24 October 2023; Issue published 17 July 2024
 View Full Text
View Full Text Download PDF
Download PDFAbstract
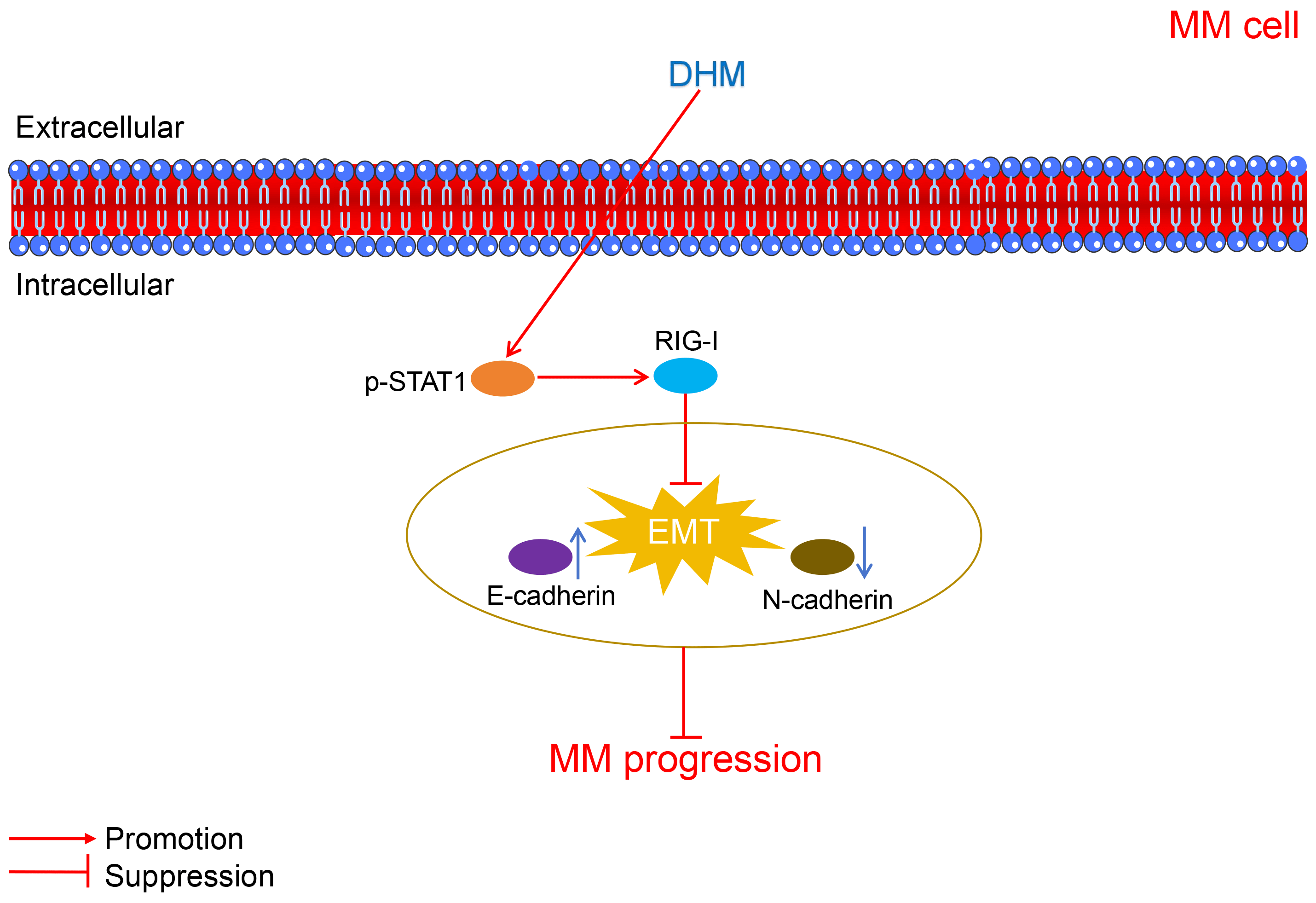
Multiple myeloma (MM) is a plasma cell malignancy and remains incurable as it lacks effective curative approaches; thus, novel therapeutic strategies are desperately needed. The study aimed to explore the therapeutic role of dihydromyricetin (DHM) in MM and explore its mechanisms. Human MM and normal plasma samples, human MM cell lines, and normal plasma cells were used for in vitro experiments. Cell counting kit-8 (CCK-8), flow cytometry, and trans-well assays were performed for the assessment of cell viability, apoptosis, migration, and invasion, respectively. Quantitative real-time polymerase chain reaction (qRT-PCR) was employed to assess the mRNA expression of signal transducer and activator of transcription 1 (STAT1) and retinoic acid-inducible gene I (RIG-I). Western blotting was employed to assess E-cadherin, N-cadherin, signal transducer, STAT1, p-STAT1, and RIG-I protein expression. A tumor xenograft model was used for in vivo experiments. Here, dihydromyricetin (DHM) dose-dependently restrained viability, apoptosis, migration, and invasion, and facilitated apoptosis of U266 cells. After DHM treatment, the E-cadherin level was increased and the N-cadherin level was decreased in U266 and RPMI-8226 cells, suggesting the inhibitory effects of DHM on epithelial-mesenchymal transition (EMT) in MM. Besides, the levels of p-STAT1/STAT1 and RIG-I were down-regulated in MM. However, the STAT1 inhibitor fludarabine undid the suppressive effect of DMH on the malignant characteristics of U266 cells. Also, DHM inhibited MM tumor growth and EMT, and activated STAT1/RIG-I pathway in vivo. Collectively, this study first revealed that DHM can restrain EMT and tumor growth in MM by activating STAT1/RIG-I signaling, which provides a novel drug for the treatment of MM.Graphic Abstract
Keywords
Cite This Article
 Copyright © 2024 The Author(s). Published by Tech Science Press.
Copyright © 2024 The Author(s). Published by Tech Science Press.This work is licensed under a Creative Commons Attribution 4.0 International License , which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
 Downloads
Downloads
 Citation Tools
Citation Tools