
 Submit a Paper
Submit a Paper Propose a Special lssue
Propose a Special lssue Open Access
Open Access
REVIEW

MicroRNAs in thyroid cancer with focus on medullary thyroid carcinoma: potential therapeutic targets and diagnostic/prognostic markers and web based tools
1 Department of Biochemistry, Faculty of Biological Sciences, North Tehran Branch, Islamic Azad University, Tehran, 1651153511, Iran
2 Department of Pathology and Laboratory Medicine, Vancouver General Hospital, Vancouver, V5Z 1M9, Canada
3 Department of Blood Cell Research, Laboratory for Red Blood Cell Diagnostics, Sanquin, Amsterdam, 1006 AN, The Netherlands
4 Cellular and Molecular Endocrine Research Center, Research Institute for Endocrine Sciences, Shahid Beheshti University of Medical Sciences, Tehran, 1985717413, Iran
* Corresponding Author: MEHDI HEDAYATI. Email:
Oncology Research 2024, 32(6), 1011-1019. https://doi.org/10.32604/or.2024.049235
Received 31 December 2023; Accepted 19 March 2024; Issue published 23 May 2024
 View Full Text
View Full Text Download PDF
Download PDFAbstract
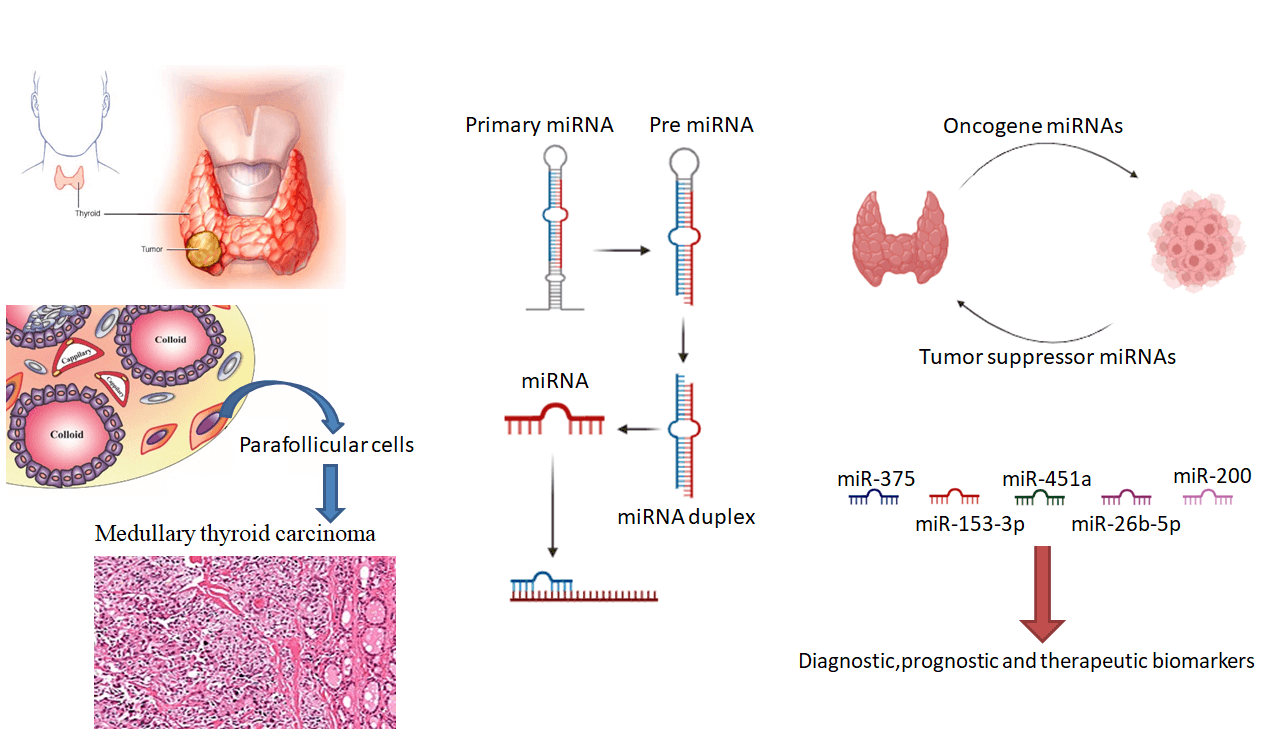
This review aimed to describe the inculpation of microRNAs (miRNAs) in thyroid cancer (TC) and its subtypes, mainly medullary thyroid carcinoma (MTC), and to outline web-based tools and databases for bioinformatics analysis of miRNAs in TC. Additionally, the capacity of miRNAs to serve as therapeutic targets and biomarkers in TC management will be discussed. This review is based on a literature search of relevant articles on the role of miRNAs in TC and its subtypes, mainly MTC. Additionally, web-based tools and databases for bioinformatics analysis of miRNAs in TC were identified and described. MiRNAs can perform as oncomiRs or antioncoges, relying on the target mRNAs they regulate. MiRNA replacement therapy using miRNA mimics or antimiRs that aim to suppress the function of certain miRNAs can be applied to correct miRNAs aberrantly expressed in diseases, particularly in cancer. MiRNAs are involved in the modulation of fundamental pathways related to cancer, resembling cell cycle checkpoints and DNA repair pathways. MiRNAs are also rather stable and can reliably be detected in different types of biological materials, rendering them favorable diagnosis and prognosis biomarkers as well. MiRNAs have emerged as promising tools for evaluating medical outcomes in TC and as possible therapeutic targets. The contribution of miRNAs in thyroid cancer, particularly MTC, is an active area of research, and the utility of web applications and databases for the biological data analysis of miRNAs in TC is becoming increasingly important.Graphic Abstract
Keywords
Supplementary Material
Supplementary Material FileCite This Article
 Copyright © 2024 The Author(s). Published by Tech Science Press.
Copyright © 2024 The Author(s). Published by Tech Science Press.This work is licensed under a Creative Commons Attribution 4.0 International License , which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
 Downloads
Downloads
 Citation Tools
Citation Tools