DOI:10.32604/phyton.2020.012476
| Phyton-International Journal of Experimental Botany DOI:10.32604/phyton.2020.012476 | |
| Article |
1Facultad de Agronomía, Universidad Autónoma de Sinaloa, Culiacán, Sinaloa, 80000, México
2Programa Regional de Posgrado en Biotecnología, Facultad de Ciencias Químico-Biológicas, Universidad Autónoma de Sinaloa, Culiacán, Sinaloa, 80030, México
3Centro de Investigación en Alimentación y Desarrollo A.C. Unidad Culiacán, Culiacán, Sinaloa, 80110, México
*Corresponding Author: Sergio Hernández-Verdugo. Email: sergioh2002mx@yahoo.com.mx
Received: 01 July 2020; Accepted: 31 August 2020
Abstract: The phenotypic variation and its distribution among species, morphotypes, and among and within populations was estimated in 71 populations pertaining to 15 morphotypes of three domesticated species of Capsicum from Mexico. Collections were made in the states of Sinaloa, Nayarit, Oaxaca, Tabasco, and Yucatán in two agroecosystems: Backyard-garden and monoculture. Fifteen phenotypic characteristics were analyzed through one-way variance analysis and multivariate analyses of principal components analysis (PCA) and hierarchical clustering using Ward’s method. The study was performed in a uniform greenhouse experiment. A high variation was found among and within populations in all the measured characteristics. Of the total variation, 13.0% was distributed among species, 27.9% among morphotypes, 8.1% among populations, and 51.0% within populations. Because plants grew in a uniform environment, these results indicate that the differences observed among and within species, morphotypes, and populations have a genetic basis. Univariate and multivariate analyses clearly differentiated morphotypes, suggesting that the category of morphotypes must be used to nominate the infraspecific variation in the domesticated Capsicum. The principal components analysis identified a total of 15 principal components that contributed to explain the total variation. The first two components explained 59.64% of the total variation and seven components explained more than 90% of the total variation. Among the measured characteristics, number of seed per fruit, weight of the fruit, width of the fruit, length of the fruit, stem diameter, days to flowering, and height of the plant contributed to component 1 variation, whereas width of the leaf, length of the leaf, and number of locules, number of fruits and number of seeds per plant contributed to component 2. The hierarchical clustering separated the populations and the morphotypes in two large different groups. One group consisted of populations collected in monoculture conditions and the other group corresponded to population collected from backyard-garden conditions. The monoculture populations were characterized mainly by their longer, wider, and higher weight fruits, plants were of less height, had smaller stem diameters, and lower number of fruits than the populations collected from the backyard-garden conditions. The backyard-garden populations of the Capsicum annuum and Capsicum frutescens species, considered wild or semi-domesticated, constituted a non-differentiated phenotypic group that does not allow dividing them in different species.
Keywords: Phenotypic variation; Capsicum annuum; Capsicum chinense; Capsicum frutescens; morphotype
 | This work is licensed under a Creative Commons Attribution 4.0 International License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. |
Mexico is one of the countries with the highest plant diversity and one of the main plants domestication centers in the world. In particular, the pepper (Genus Capsicum) was one of the first plants domesticated in the American continent [1]. The pepper is one of the domesticated plants of high economic and nutritional value worldwide. The genus Capsicum is native of South America and is constituted by approximately 31 species distributed from the South of the USA to the North of Argentina, with a number of basic chromosomes that can be n = 12 or n = 13 [2–4]. Of this genus, the following C. annuum, C. chinense, C. frutescens, C. pubescen, and C. baccatum have been domesticated. In Mexico, the first four of these species are being cultivated [3–4]. Species C. annuum, C. chinense, and C. frutescens are tightly related and constitute the C. annuum-C. chinense-C. frutescens known complex [5].
Of the domesticated species, C. annuum is the one of greatest economic relevance, it is widely cultivated in the world and it was domesticated in Mexico [5,6]. This species presents a great variation in size, shape, and color of the fruits and it is cultivated in all agricultural regions of Mexico; this species comprises the peppers known as “Cola de rata” or “de árbol”, “Jalapeños”, and “Cascabel”, among others. Their wild populations are widely distributed from the South of the USA to the low-altitude regions of Peru [5,3]. The C. chinense species is mainly cultivated in South America. Its most representative type in Mexico is the “Habanero” pepper, which is grown in Campeche and Yucatan. Its wild form is found in Peru et al. [6]. The C. frutescens species is cultivated in tropical and subtropical regions of Mexico. To this species pertains the “Tabasco” pepper, from which the well-known “Tabasco” sauces are made. Another type of peppers of this species is the “Pico de paloma” morphotype, which is found in backyard conditions in the houses of the rural regions of the state of Tabasco [7–11]. The wild materials of C. frutescens are distributed in the Center and Southeast of Mexico until Colombia. It is found in secondary vegetations and in untouched places at low altitude. It can grow as a herbaceous plant or as a shrub capable of climbing. It can reach 2 m in height [3]. It is considered that this species was domesticated in Mexico [12].
Domestication is an evolutionary process that operates under the influence of human activities. During this process, cultivated plants have developed characteristics that result from adaptations to the local ecological conditions, to cultivating conditions, and to the needs of humans. During the course of domestication, in the species of cultivated plants occur differences among the domesticated populations of the same species and within them and their closest wild relatives. To most domesticated species, the pepper (Capsicum) presents intraspecific variations, particularly, in the part of the plant used by humans, in this case, the fruits [3,4].
In Mexico, since pre-Columbian times, the indigenous communities have selected the characteristics of greatest human interest, such as the size and shape of the fruits. This activity performed for thousands of years has produced a high morphological variability and many cultivars adapted to local conditions, with stable yields under a traditional management system.
Since it has been recognized that the evolutionary potential of a species depends on its available genetic variability, one of the core tasks in evolutionary biology, as well as in genetic improvement, and the collection and maintenance of the vegetal genetic resources is to estimate the amount of genetic variability maintained in the species of domesticated plants, their races or local varieties, wild relatives, and the pattern of distribution of such variation [13].
Studies with isoenzymes, molecular markers RAPDs, and microsatellites [14–17] have indicated that the populations of wild and domestic peppers of the Northwest of Mexico maintain high levels of genetic variations within and among their populations. However, the amount of variation in quantitative characters and their distribution among and within species, morphotypes, and population are not known yet. In this study, 71 populations of 15 morphotypes of the C. annuum, C. chinense, and C. frutescens species of different regions of Mexico were analyzed in a uniform greenhouse experiment, aiming at estimating the phenotypic variations and their relative distribution among and within species, morphotypes, and populations.
2.1 Plant Material and Collection Sites
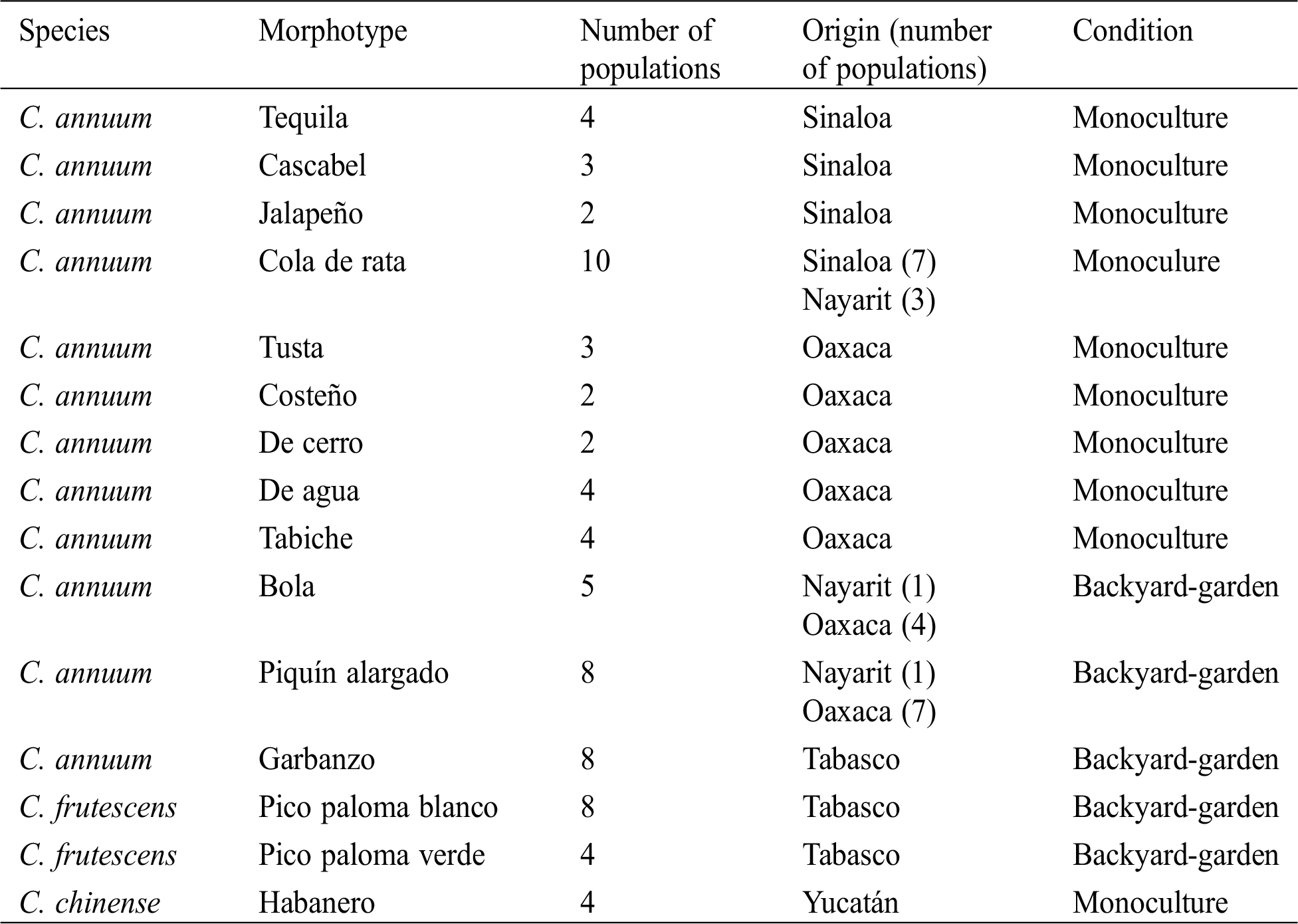
Ripe fruits of 71 pepper populations were collected in two agroecosystems: backyard-garden and monoculture. The backyard or garden lies near the houses in rural areas and their production is generally for self-consumption. Monoculture is performed with modern technology and the production is meant for local or national marketing. The 71 collected populations pertained to 15 morphotypes of the C. annuum, C. chinense, and C. frutescens species (Tab. 1 and Fig. 1). The names of the morphotypes correspond to those used by the inhabitants of the rural area from which they were collected. Seeds were collected in the states of Sinaloa, Nayarit, Oaxaca, Tabasco, and Yucatan across Mexico (Fig. 1).
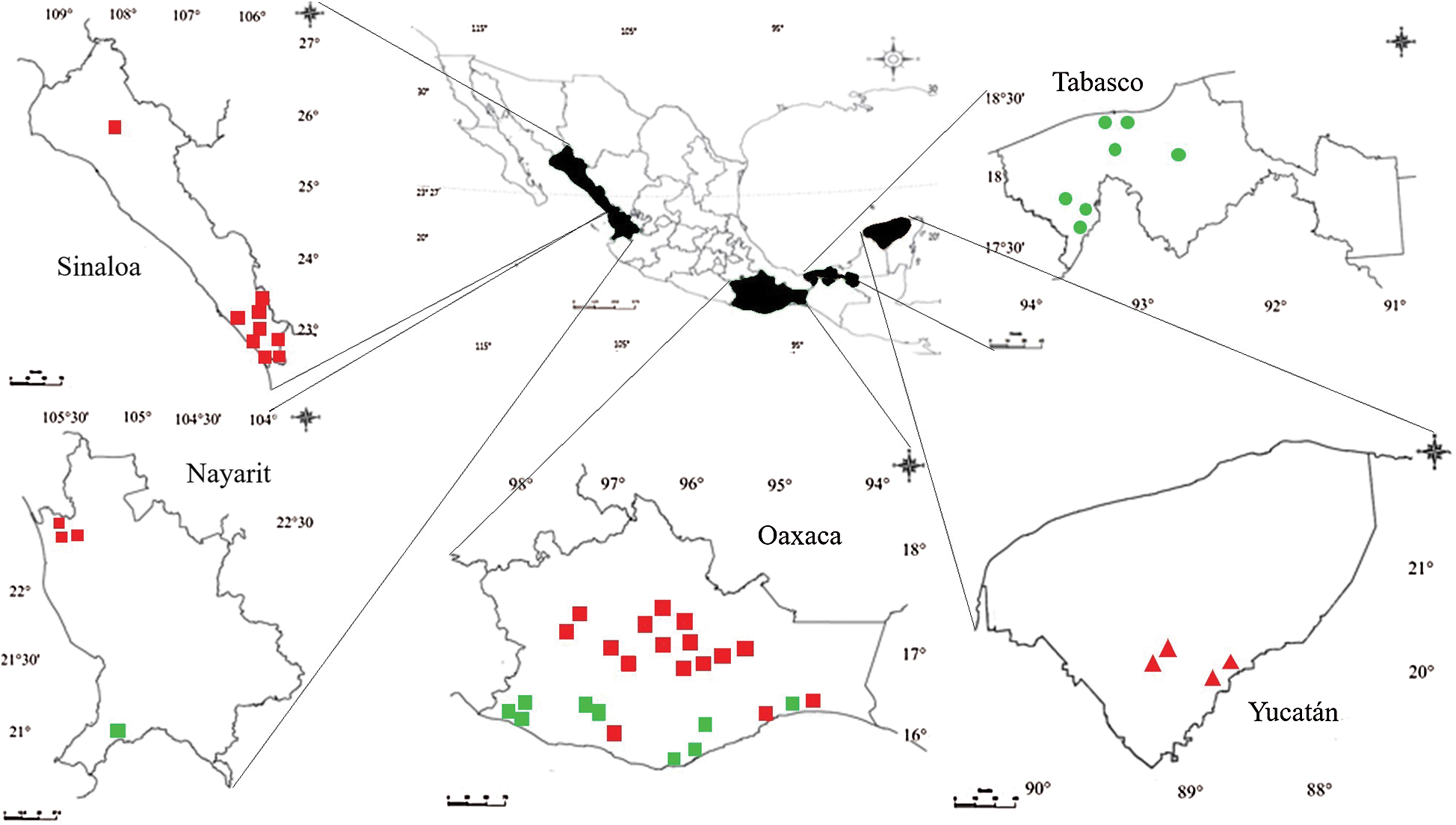
Figure 1: Locations of the 71 populations of Capsicum annuum, Capsicum frutescens, and Capsicum chinense collected in the states of Sinaloa, Nayarit, Oaxaca, Tabasco, and Yucatán across Mexico (  C. annuum monoculture;
C. annuum monoculture;  C. annuum backyard-garden;
C. annuum backyard-garden;  C. chinense monoculture;
C. chinense monoculture;  C. frutescens backyard)
C. frutescens backyard)
Table 1: Species, morphotypes, number of populations and origin (number of populations) of the studied pepper (Capsicum) species
After the collection, 100 seeds of each of the 71 populations were soaked in a solution with 1000 ppm of gibberellic acid (GA) for 36 h. Afterwards, the GA-treated seeds were left to germinate in 128-wells polystyrene plates with Peat-Moss as substrate; 75 days after sowing, six plants were taken from each population and individually transplanted into polystyrene bags containing 10 kg of alluvium soil, at a separation of 30 cm per bag, 1.30 m between furrows, with drip irrigation, in greenhouse conditions under a shadow-mesh that allowed 75% of sun light. The experiment was conducted in completely randomized design with six repetitions. The experimental unit corresponded to one plant.
Six months after transplanting, their characteristics were measured: (1) days to flowering, (2) days to fruit ripening, (3) height of the plant (cm), (4) diameter of the stem (mm), (5) length and (6) width of the leaf (mm), (7) length of the pedicel of the fruit (mm), (8) number of fruits per plant, (9) length and (10) width of the fruit (mm), (11) weight of the fruit (g), (12) number of locules of the fruit, (13) number of seed per fruit, (14) individual weight of the seed (mg), and (15) number of seed per plant. The length, width, weight, and number of locules of the fruit were obtained from a sample of five fruits per plant. The weight of the seed was obtained by weighing the seed of each of the five fruits and dividing by the number of seed per fruit. The number of seed per plant was obtined by multipiying the number seed per fruit with the number of fruits per plant. Fruits and seeds were weighed in an analytical balance model TP-214 (Denver-Instrument, Co., Bohemia, NY, USA), with an accuracy of 0.1 mg.
Descriptive analysis was applied to estimate the phenotypic variability of all collections. The mean, minimum and maximum values, standard deviation (SD) and the coefficient of variation (CV) of the measured characteristics were estimated. The CV is the standar deviation expresed as a percentage of the mean. The CV is independent of the unit of measurement and is experessed as a percentage. A nested-variance analysis was performed to determine the distribution of the relative amounts of variation among species, among morphotypes within species, and among populations within the morphotypes for each measured phenotypic characteristic. The variation within populations was estimated through the residual term of the nested-variance analysis. Species, morphotypes within species, and populations within morphotypes were considered random factors. A one-way variance analysis was performed to determine the differences among morphotypes. The means of populations were used to perform multivariate analysis based on principal components analysis (PCA) and hierarchical clustering with the Ward’s method. Eigenvectors with an associated eigenvalue higher than 1.0, or explaining >10% of the total variance, were considered significant [18]. Sets of covarying characters were identified by the relative loadings of the traits on the significant principal components [19]. Scatterplot of the positions of the centroids corresponding to each population in the multivariate space defined by the first and second components was obtained to illustrate inter and intra-Morphotype variation. All analyses were performed with the JMP statistical software [20].
Results of the descriptive analysis revealed a high variation for most of the measured characteristics of the studied Capsicum populations (Tab. 2).
Table 2: Mean, standard deviation (SD), minumun and maxium values, and coefficient of variation (CV) of 15 characteristics measured in 71 populations of C. annuum, C. frutescens, and C. chinense species of Mexico
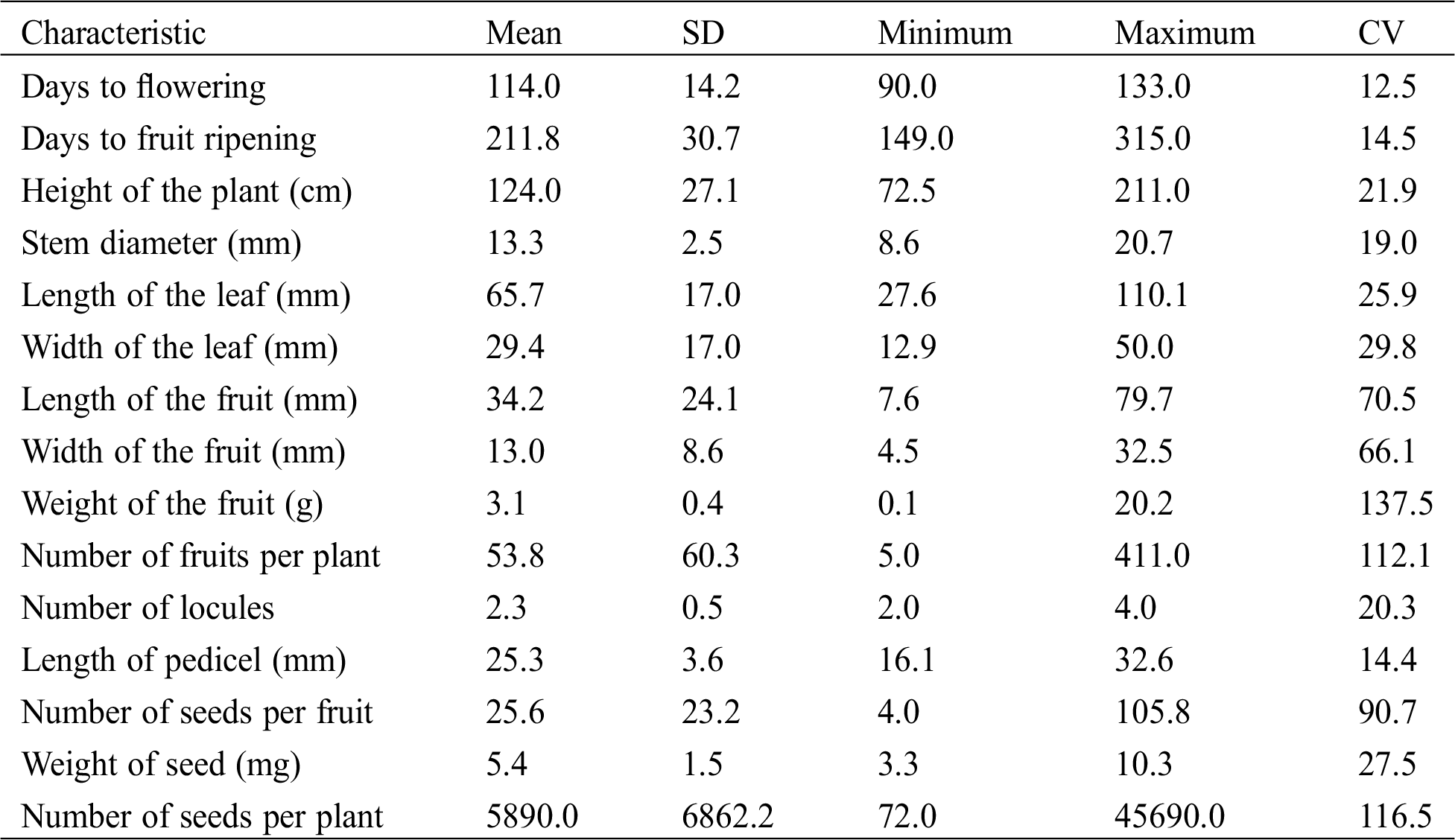
Days to flowering and days to fruit ripening varied from 90.0 to 133.0 and 149.0 to 315.0, respectively. The height of the plant varied from 72.5 to 211.0 cm, with a mean of 124 cm. The length and width of the leaf varied from 27.6 to 110.1 and 12.9 to 50 mm, respectively. The length of fruits varied from 7.6 to 79.7 mm, with a mean of 34.2 mm, whereas the width of fruits varied from 4.5 to 22.5 mm, with a mean of 13.0 mm. The weight of the fruit varied widely, from 0.1 to 20.2 g. The number of fruits per plant varied widely, from 5 to 411. The number of seed per fruit varied from 4 to 106. The number of seeds per plant also varied widely, from 72 to 45,690 (Tab. 2).
The characteristics corresponding to weight of fruit, number of fruits, and number of seeds per plant presented a CV higher than 110%, whereas the width and length of the fruit and number of seeds per fruit had a CV close or higher than 70%. Only the characteristics concerning days to flowering, days to ripening of fruits, and length of the pedicel showed a CV lower than 15% (Tab. 2). The average CV of the 15 characteristics was of 51.9%.
3.2 Distribution of the Variation
The nested-variance analyses revealed that in average, 13% of the total phenotypic variation observed in the measured characteristics was distributed among species, 27.9% among morphotypes, 8.1% among populations, and 51.0% within populations (Tab. 3). All the traits, except height of the plant, length of the leaf, length of the fruit, weight of the fruit, length of pedicel and number of seeds per plant, showed a significantvariation among the species (Tab. 3). Differences among species went from 0% (length of leaf, length of fruit, weight of fruit, length of pedicel, and number of seeds per plant) to 45.1% (number of locules). The characteristic width of the leaf presented a value greater than 30%, whereas the characteristics stem diameter, width of fruit, days to flowering, and number of seeds per fruit showed a higher than 20% differentiation among species (Tab. 3).
Table 3: Percentages of phenotypic variation among species, morphotypes, populations, and within the population (residual) measured in 71 populations of 15 morphotypes pertaining to three pepper (Capsicum) species of Mexico
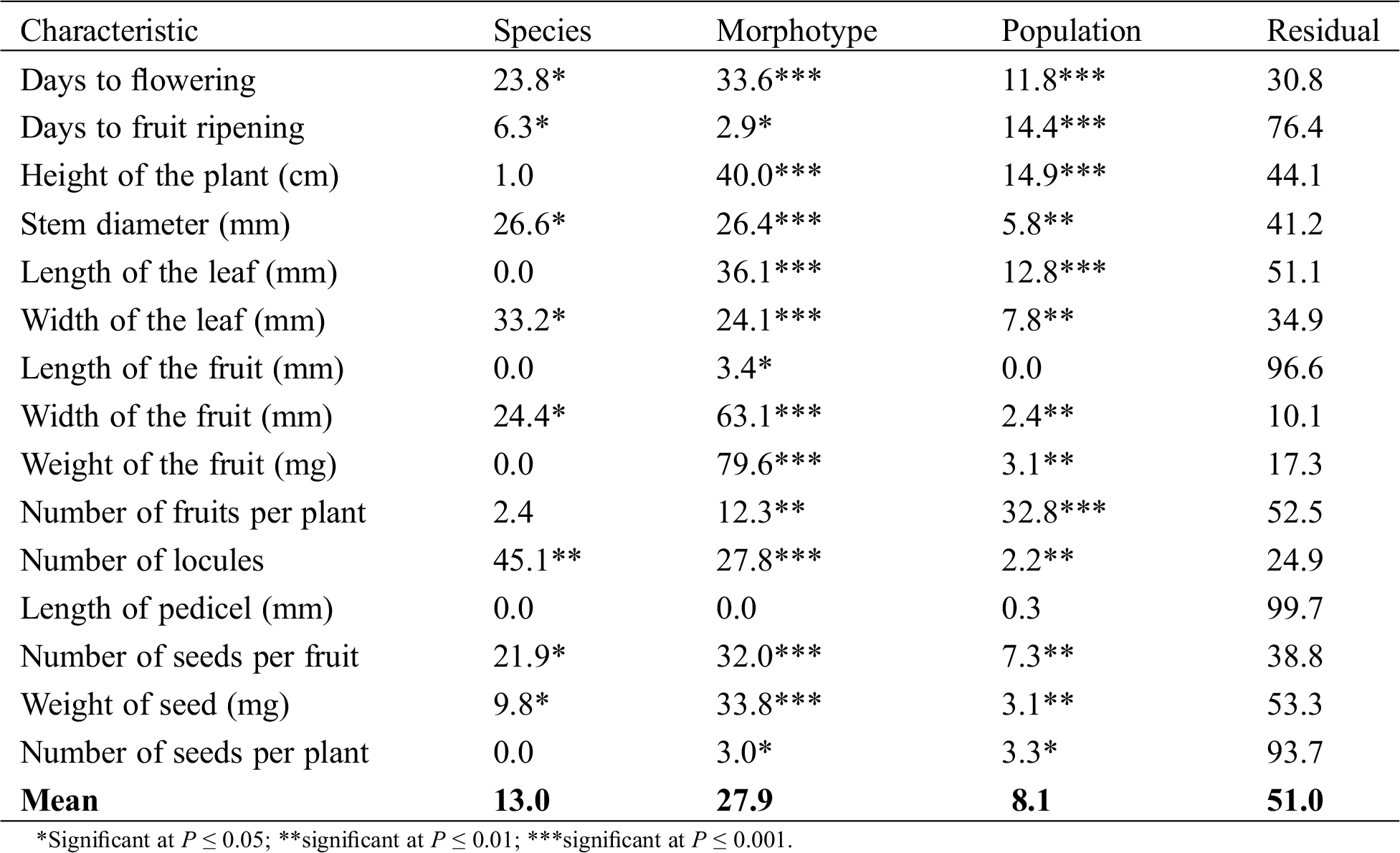
All the traits, except length of pedicel, showed a significant variation among the morphotypes (Tab. 3). The relative amount of variation among morphotypes ranged from 0% (length of pedicel) to 79.6% (weight of the fruit) and 63.1% (width of fruit) of the total variation. The characteristic height of plant presented 40% variation among morphotypes, whereas the length of the leaf, weight of seed, number of seeds per fruit, and days to flowering presented more than 30% differences among morphotypes. The characteristics number of locules per fruit and stem diameter showed differences higher than 25% (Tab. 3).
All the traits, except length of the fruit and length of pedicel, showed a significant variation among the populations (Tab. 3). Variation among populations was relatively low in all characteristics, except in number of fruits per plant, which presented a higher than 30% variation (Tab. 3). In contrast, the variation observed within populations was high in most of the characteristics. Only the width of the fruit presented a 10.1% variation (Tab. 3). The remainder of the characteristics presented variation percentages from 17.3% (weight of the fruit) to 99.7% (length of pedicel) and 96.6 % (length of fruit).
3.3 Differentiation Among and Within Species and Morphotypes
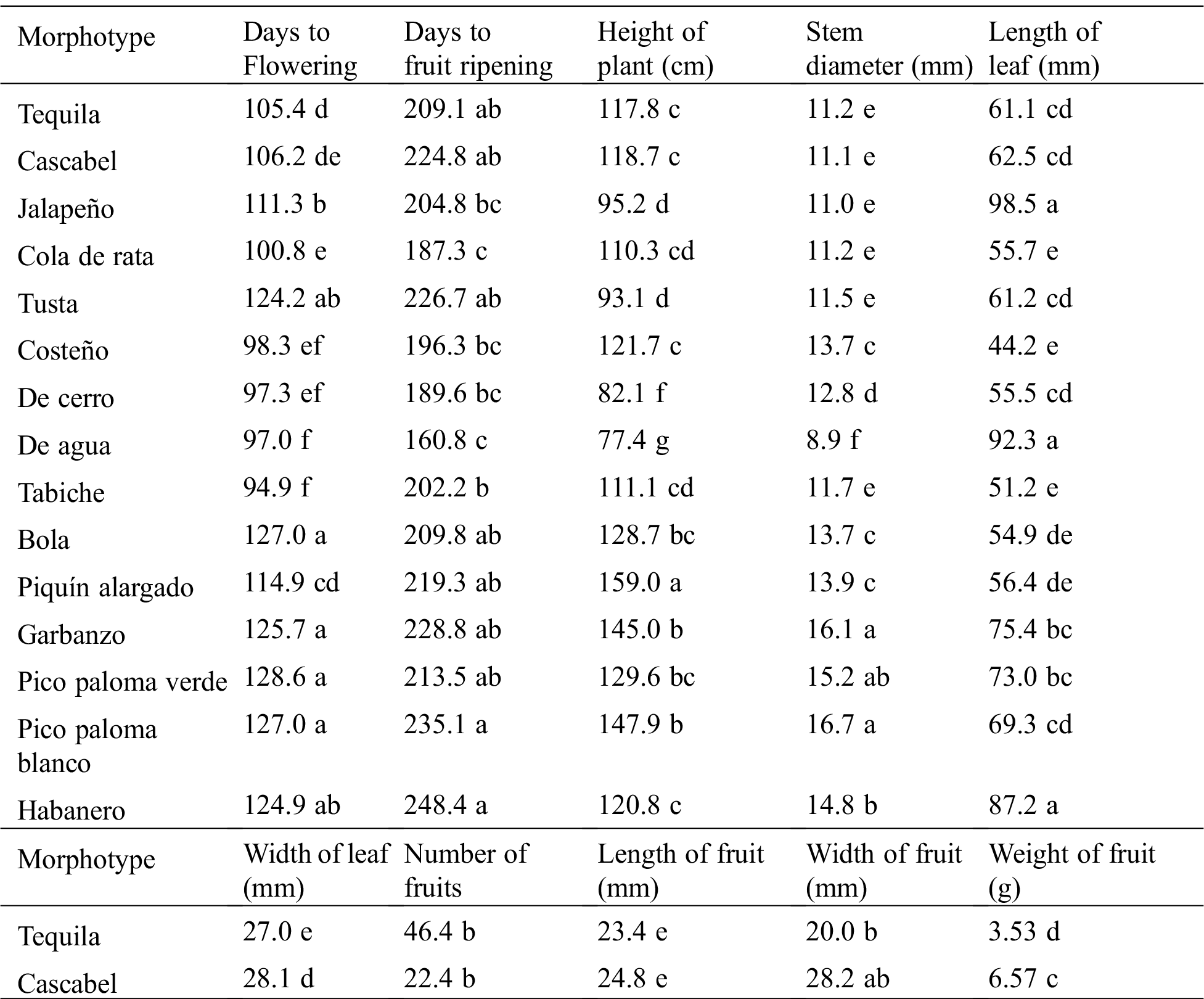
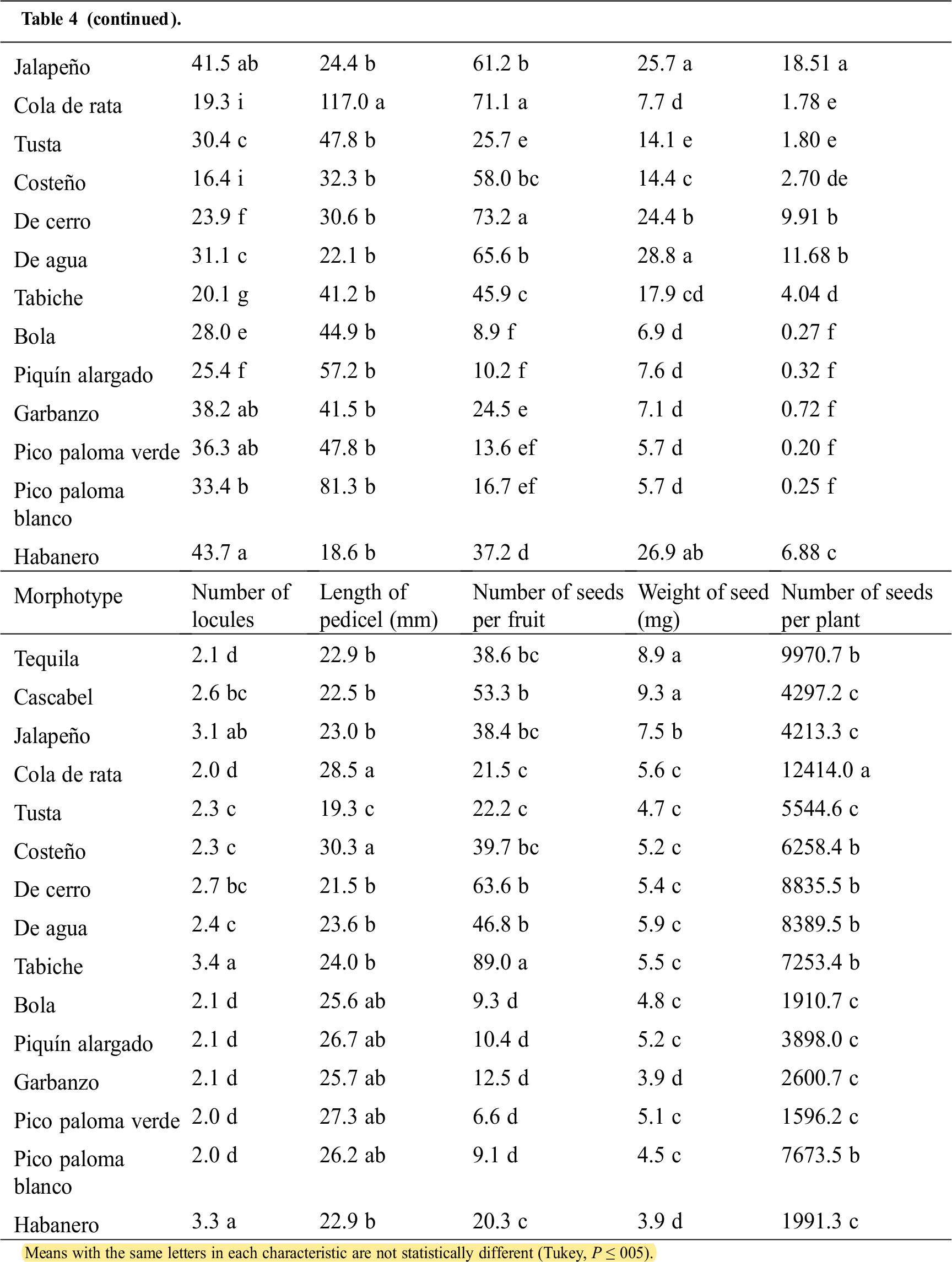
The one-way variance analysis revealed that the Capsicum morphotypes differed significantly in all the assessed characteristics (Tab. 4).
Table 4: Means of characteristics measured in 15 Capsicum morphotypes
In the principal component analysis, a total of 15 principal components were identified that contributed to the total variation (Tab. 5). The first two components explained 59.64% of the total variation, and the first seven components explained more than 90% of the total variation (Tab. 5).
Table 5: Eigenvalues and proportion of total variance explained by each principal components
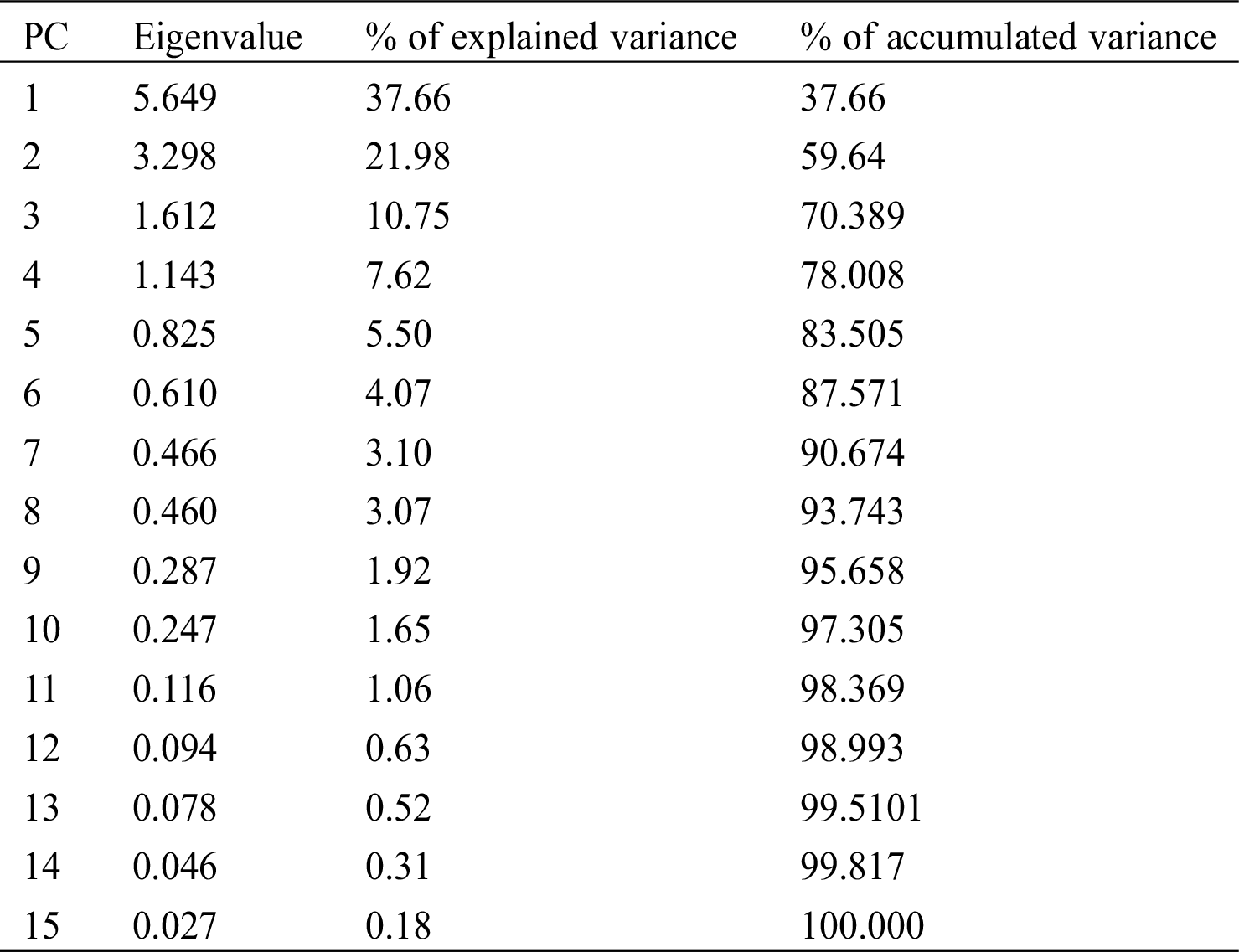
The first component (PC1) explained 37.66% of the variations (Tab. 5), which was contributed by seven strongly, correlated characteristics (Tab. 6). Positively correlated traits that contributed to the total variation of PC1 were related to number of seed per fruit, weight of the fruit, width of the fruit, and length of the fruit (Tab. 6). Negatively correlated traits that contributed to the variation of the first component were related to stem diameter, days to flowering, and height of the plant (Tab. 6). The second component (CP2) explained 21.98% of the variations (Tab. 5) and was mostly defined by the characteristics width of the leaf, length of the leaf, and number of locules, positively correlated (Tab. 6). The characteristics number of fruits and number of seeds per plant negatively correlated. The third component (CP3) explained 10.75% of the variation (Tab. 5) and was determined by the characteristics number of fruits per plant, number of seed per plant, length of the leaf, and width of the leaf positively correlated (Tab. 6).
Table 6: Results of the principal components analysis performed on 15 phenotypic characteristics in 71 populations of C. annuum, C. Chinense, and C. frutescens species of Mexico. Variables with the highest weight (>10% of the variance of each principal component) are indicated in bold
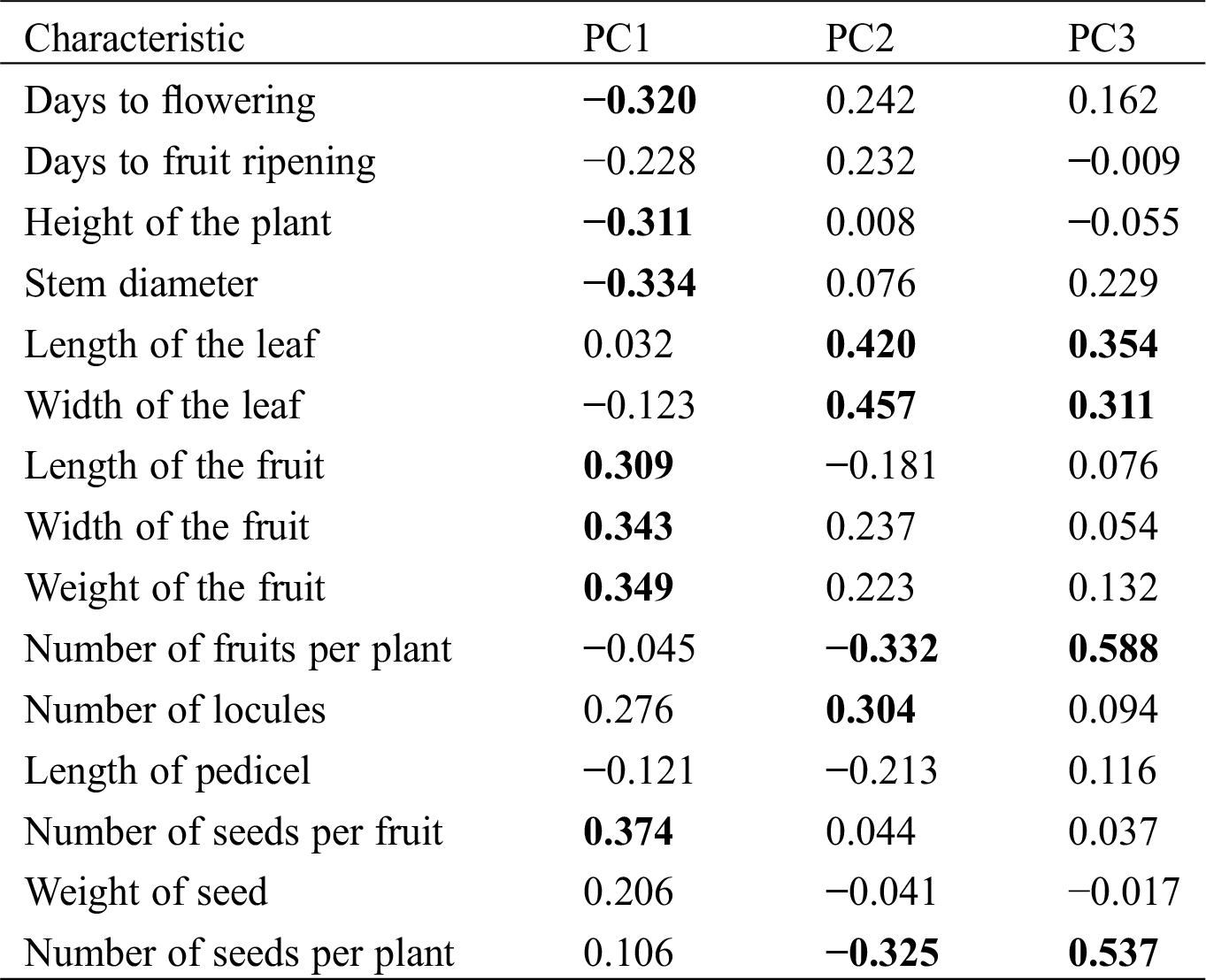
Populations and morphotypes were differentiated in the two-dimensional space of the principal components 1 and 2 (Fig. 2). The principal component 1 distinguished the populations “Pico de paloma blanco” y “Pico de paloma verde” (C. frutescens), “Piquín alargado” (C. annuum), together with all the populations of “Garbanzo” (C. annuum) and “Bola” (C. annuum) in the region of lower length, width and weight of fruits, and less number of seeds per fruit, but with plants of more height, greater stem diameter, and longer times to flowering. Whereas the four populations of the “De agua” morphotype, the two “Jalapeños”, and the two “De cerro” morphotypes occupied the opposite region. The remainder populations of the “Cascabel”, “Tabiche”, “Tequila”, “Tusta”, “Costeño”, and “Cola de rata” morphotype were grouped in the central region.
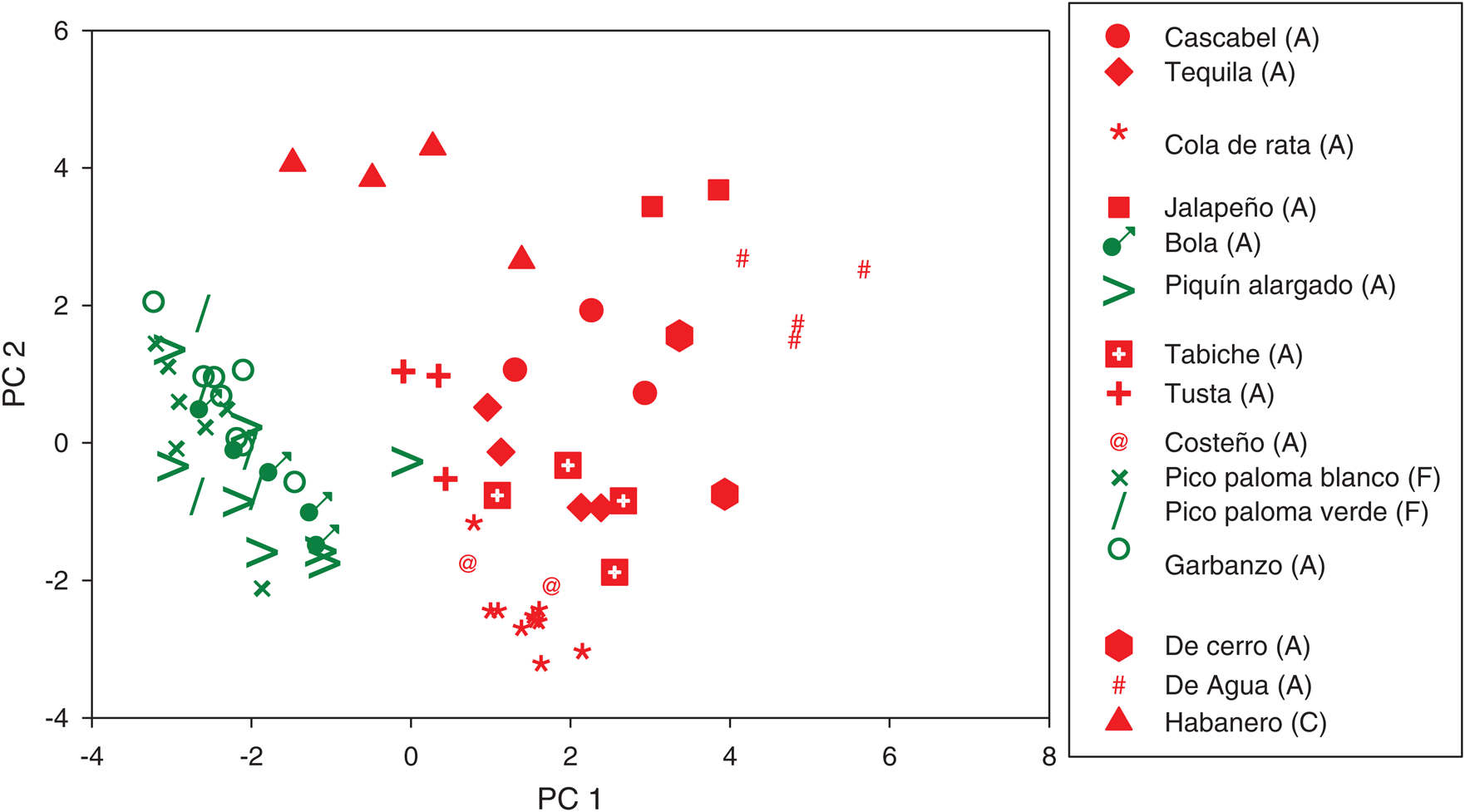
Figure 2: Plot of principal components 1 and 2 for the 71 populations of 15 morphotypes of three pepper species from Mexico. C. annuum (A); C. frutescens (F); C. chinense (C) (Green = Backyard-garden; Red = Monoculture)
The principal component 2 separated the populations of the “Habanero” (C. chinense) morphotype in the region of wider and longer leaves, greater number of locules per fruit, but less number of fruits and less seeds per plant. The populations “Cola de rata” occupied the opposite region. The other populations were distributed in the intermediate region.
The dendrogram constructed with the Ward’s method allowed distinguishing most of the studied morphotypes in two large groups (Fig. 3). In the first group were all the pepper populations collected in monoculture conditions, as well as three populations of “Piquín alargado” and one population of “Pico de paloma blanco”. The second group corresponded to the populations collected from backyard-garden conditions (Fig. 3). The monoculture populations were characterized for having longer, wider, heavier fruits (Fig. 4), plants of lesser height, stem diameter, less time to flowering, and smaller number of fruits. Whereas, the backyard-garden populations showed smaller and lower weight fruits (Fig. 4). The plants were higher, had a greater stem diameter, longer times to flowering, and a greater amount of fruits.
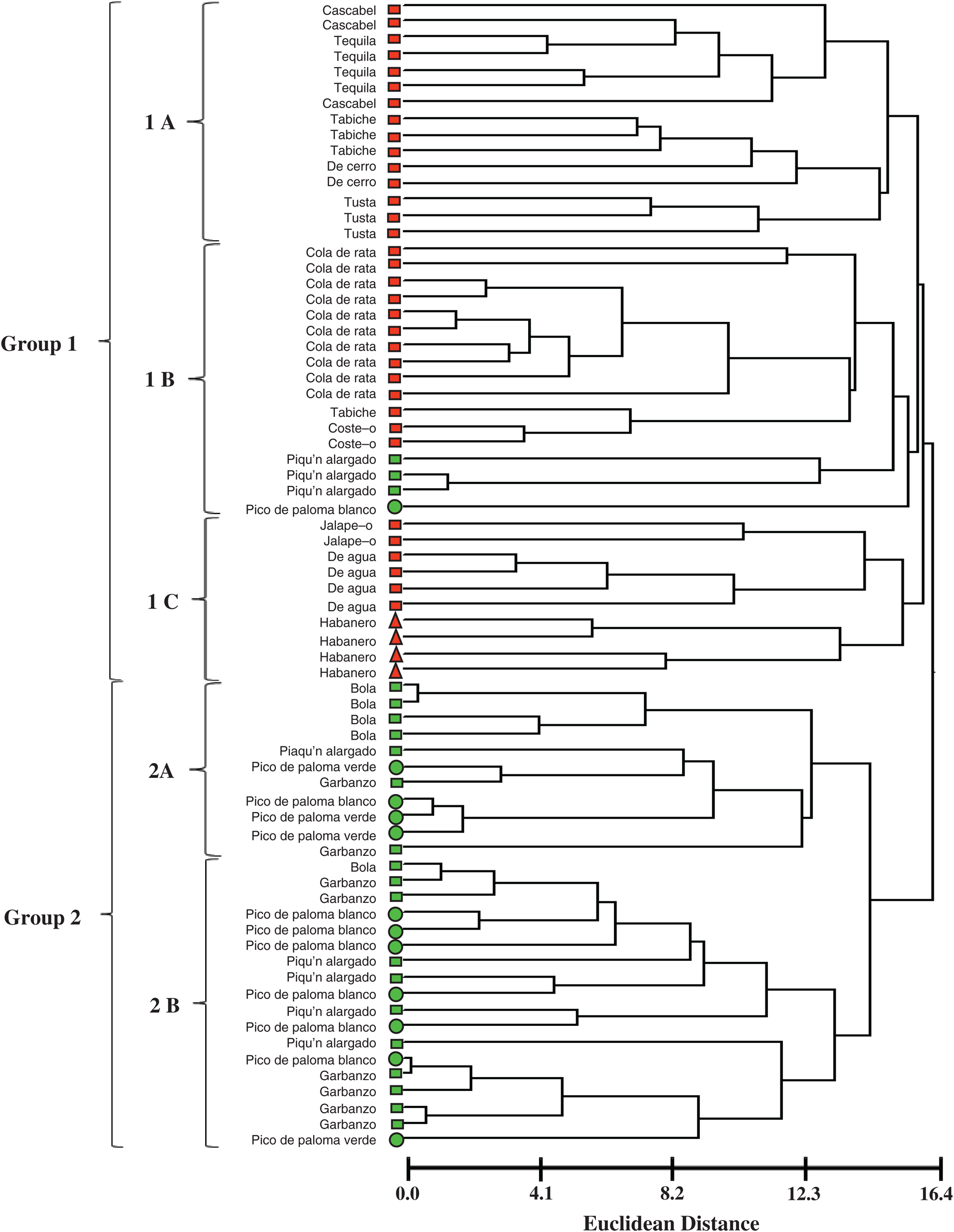
Figure 3: Dendrogram with the Ward’s clusters analysis of the 71 populations pertaining to 15 morphotypes of C. annuum, C. frutescens, and C. chinense species from Mexico (  C. annuum monoculture;
C. annuum monoculture;  C. annuum backyard-garden;
C. annuum backyard-garden;  C. chinense monoculture;
C. chinense monoculture;  C. frutescens backyard-garden)
C. frutescens backyard-garden)

Figure 4: Representative fruit variability of different chili pepper (Capsicum spp.) morphotypes allocated in the dendrogram groups
Three subgroups were formed within the first group. The first subgroup (1A) was constituted by three populations of the “Cascabel” morphotype and the four “Tequila” populations, characterized mainly by their globous fruits. These populations were related with three of the four “Tabiche” morphotype populations, the two “De cerro” populations, and the three populations of the “Tusta” morphotype.
The second subgroup was constituted by 17 populations (1B). Here are the 10 populations of the “Cola de rata” morphotypes, related with one “Tabiche” population, the two “Costeño” morphotype populations, three of the eight “Piquín alargado” populations, and one “Pico de paloma blanco”.
The third subgroup was constituted by 10 populations (1C). In this group comprised the “Jalapeño” morphotype populations, the four “De Agua” populations, and the four “Habanero” (Capsicum chinense) populations. The populations of each one of these morphotypes were grouped together and were differentiated from the other morphotypes.
The second group was composed of 29 populations, all collected in backyard conditions (Fig. 3). These morphotypes present smaller fruits (Fig. 4) and have a lower domestication level. This group is constituted by two subgroups. The “Bola” (C. annuum), “Piquín alargado” (C. annuum), “Garbanzo” (C. annuum), “Pico de paloma blanco” and “Pico de paloma verde” (C. frutescens) are distributed in both subgroups.
Of the 15 measured characteristics, 12 had a CV higher than 20%, indicating high levels of phenotypic variation in the studied populations. It is considered that a CV higher than 20% means the presence of high variation in plant populations or species useful for agriculture and food [21–22]. In this sense, it has been interpreted that a CV above 20% as the presence of a wide variation within populations and species of the Cola [21], Pelargonium [22], Helianthus [23] genera. In Capsicum, has been reported a CV higher than 20% in six of nine quantitative characteristics measured in 19 wild populations of pepper in the Northwest of Mexico [24]. Another study in Capsicum [25] reported that of 18 measured characteristics, 13 presented a CV higher than 20% in 148 domesticated populations of C. annuum, C. frutescens, and C. chinense.
The characteristics related with the fruits presented the highest CVs, followed by the vegetative characteristics and the days to flowering and ripening of the fruits. The CVs of the characteristics related to the fruits ranged from 66.1% (width of fruit) to 137.5% (weight of fruit). The CVs of vegetative characteristics varied from 19.0% (stem diameter) to 29.8% (width of leaf). In contrast, the characteristics of days to flowering and to ripening of fruits presented the lowest CVs. These results are in line with those obtained by Orobiyi et al. [25], who, in a study with 148 populations of the C. annum C. frutescens and C. chinense species, found that the variation in quantitative characters is higher in those related to the fruits, followed by the vegetative characteristics and days to flowering and ripening of fruits. These results agree with those obtained by Villota-Cerón et al. [26] and Castañón Nájera et al. [8], who, in studies of 68 and 40 accessions, respectively, pertaining to the three Capsicum species studied herein, reported that the CVs were higher for the fruit characteristics, followed by the vegetative. This higher variation observed in the characteristics related with the fruits can be due to the artificial selection exerted by humans on the characteristic most relevant in terms of nutritional and economic interest in cultivated Capsicum. These results show that the studied Capsicum populations maintain higher levels of variation, which could be used for genetic improvement programs.
Of the total phenotypic variation, 13.0% was distributed among species, 27.9% among morphotypes, 8.1% among populations, and 51.0% within populations. Because plants grew in a uniform environment, these results indicate that the variation observed among and within species, morphotypes, and populations can be considered a genetic variation among and within them. These results agree with those obtained with molecular markers of isoenzymes [14], RAPDs [16], microsatellites [17], and in quantitative characteristics [24,27,28], which have indicated that the wild and domesticated Capsicum species maintain a higher genetic variation within and among populations. According to the results of this study, the strategy to identify the genetic diversity in these species consists in emphasizing the collection of a significant number of individual plants from each population.
Whthin most cultivated species there is a considerable amount of hierarchical structured variation. One problem in the taxonomic treatment of a genus containig both cultivated and wild plants is how treat infraespecific variation, particulary in the cultivated taxa [29]. In this study, the differences among morphotypes explained a good amount of the total phenotypic variation (27.9%). This high variation among morphotypes indicates that the morphotype category should be considered for the infraspecific classification in the domesticated Capsicum. These results indicate that the names given by the farmers to the different morphotypes has a biological basis. The high variation among morphotypes regarding the characteristics related with the fruits indicates that farmers, through artificial selection, have set the desired characteristics of each morphotype for their use and particular consumption. Morphotype category make it posible to distingish the variation above the cultivar level but below the level species [29].
4.3 Differentiation Among and Within Species and Morphotypes
The Capsicum morphotypes showed significant differences in all the assessed characteristics (Tab. 4), indicating that the category of morphotype is valid to discern the infraspecific variation in the cultivated Capsicum species. These results agree with those obtained by Castellón-Martínez et al. [9], who reported significant differences in all the characteristics measured in the plant and fruits of “De agua”, “Nanche”, “Piquín”, “Solterito”, “Tabaquero”, and “Tusta” pepper morphotypes collected in the state of Oaxaca, Mexico.
The multivariate analysis of principal components distinguished the populations of the “Habanero” (C. chinense) morphotype from other populations. “Habanero” populations were characterized by having plants with wider and longer leaves, larger number of locules per fruit, less number of fruits, and less amount of seeds per plant. This result agrees with those of Castañón-Nájera et al. [7], who, in a study of 11 morphotypes from the state of Tabasco, Mexico, reported that the Habanero morphotype differentiated from the rest of morphotypes, including the “Pico de paloma” (C. frutescens) and “Garbanzo” (C. annuum) morphotypes, analyzed in this study.
Principal component analysis and hierarchical clustering with the Ward’s method separated the collected populations in monoculture conditions from those of backyard-garden conditions. Monoculture populations were characterized by longer, wider, and heavier fruits, but plants of less height, smaller stem diameter, and less number of fruits than the populations collected from backyard-garden conditions. These results agree with those of Castañón-Nájera et al. [7], who, in a collecting and characterization study of “Garbanzo” (C. annum), “Pico de paloma” (C. frutescens), “Ojo de Cangrejo” (C. annum), “Amashito” (C. annum), “Corazón de pollo” (C. annum), and “Habanero” (C. chinense) morphotypes, reported that the wild or semi-domesticated populations constituted a different group from the commercial populations.
Within the group of monoculture populations, most populations of each morphotype grouped together, confirming that the category of morphotype is useful to distinguish the infraspecific variation in the domesticated Capsicum.
The four populations of “Habanero” pepper (C. chinense) grouped together and were related with the populations of the “De agua” (C. annuum) and “Jalapeño” (C. annuum) morphotypes.
The populations of the backyard morphotypes considered as wild or semi-domesticated, characterized mainly by their small fruits and used for self-consumption or to be sold in local markets, constituted a different group from the monoculture populations. This group comprises the populations of “Bola” (C. annuum), “Piquín alargado” (C. annuum), “Garbanzo” (C. annuum), “Pico de paloma verde” and “Pico de paloma blanco” (C. frutescens) morphotypes distributed in two non-differentiated phenotypic subgroups. These results agree with Pickersgil et al. [30], who reported that the wild forms of the C. annuum, C. frutescens, and C. chinense species form a poorly differentiated complex that does not allow dividing them in distinct species; the results also agree with those of Villota-Cerón et al. [26] and Palacios-Castro et al. [31] who, in studies of morphological characterizations, were unable to discriminate among the C. annuum, C. frutescens, and C. chinense species, and concluded that the three constitute one morphological group.
Funding Statement: This research was funded by the Universidad Autónoma de Sinaloa (Project PROFAPI 2015/123).
Conflicts of Interest: The authors declare that they have no conflicts of interest to report regarding the present study.
References
1. MacNeish, R. S. (1964). Ancient Mesoamerican civilization. Science, 143(3606), 531–537. DOI 10.1126/science.143.3606.531.
2. Carrizo-García, C., Barfuss, M. H. J., Sehr, E. M., Barboza, G. E., Samuel, R. et al. (2016). Phylogenetic relationships, diversification and expansion of chili peppers (Capsicum, Solanaceae). Annals of Botany, 118(1), 35–51. DOI 10.1093/aob/mcw079. [Google Scholar] [CrossRef]
3. Hernández-Verdugo, S., Dávila-Aranda, P., Oyama, K. (1999). Síntesis del conocimiento taxonómico, origen y domesticación del género Capsicum. Boletín de la Sociedad Botánica de México, 64, 65–84. DOI 10.17129/botsci.1583. [Google Scholar] [CrossRef]
4. Hernández-Verdugo, S. (2018). El chile silvestre ecología, evolución y genética. Colegio de Postgraduados, Texcoco, Estado de México, México. Universidad Autónoma de Sinaloa, Culiacán, Sinaloa, México. [Google Scholar]
5. Pickersgill, B. (1971). Relationships between weedy and cultivated forms in some species of chili peppers (genus Capsicum). Evolution, 25(4), 683–691. DOI 10.2307/2406949. [Google Scholar] [CrossRef]
6. Pickersgill, B. (1984). Migration of chili peppers, Capsicum spp. in the Americas. In: Stone, D. (ed.Pre-columbian plant migration, pp. 105–123. Cambridge, USA: Harvard University Press. [Google Scholar]
7. Castañón-Nájera, G., Latournerie-Moreno, L., Mendoza-Elos, M., Vargas-López, A., Cárdenas-Morales, H. (2008). Sampling and characterization of pepper chilli (Capsicum spp) in Tabasco, Mexico. Phyton-International Journal of Experimental Botany, 77, 189–202. [Google Scholar]
8. Castañón-Nájera, G., Latournerie-Moreno, L., Lesher-Gordillo, J. M., De la Cruz-Lázaro, E., Mendoza-Elos, M. (2010). Identificación de variables para caracterizar morfológicamente colectas de chile (Capsicum spp.) en Tabasco, México. Universidad y Ciencia, 26(3), 225–234. [Google Scholar]
9. Castellón-Martínez, E., Carrillo-Rodríguez, J. C., Chávez-Servia, J. L., Vera-Guzmán, A. M. (2014). Phenotype variation of chile morphotypes (Capsicum annuum L.) native to Oaxaca, Mexico. Phyton-International Journal of Experimental Botany, 83, 225–236. [Google Scholar]
10. Nárez-Jiménez, C. A., De la Cruz-Lázaro, E., Gómez-Vázquez, A., Castañón-Nájera, G., Cruz-Hernández, A. et al. (2014). La diversidad morfológica in situ de chiles silvestres (Capsicum spp.) de Tabasco, México. Revista Fitotecnia Mexicana, 37(3), 209–215. DOI 10.35196/rfm.2014.3.209.
11. De la Cruz-Lázaro, E., Márquez-Quiroz, C., Osorio-Osorio, R., Preciado-Rangel, P., Márquez-Hernández, C. (2017). Caracterización morfológica in situ de chile silvestre Pico de paloma (Capsicum frutescens) en Tabasco, México. Acta Universitaria, 27(2), 10–16. DOI 10.15174/au.2017.1083. [Google Scholar] [CrossRef]
12. Loaiza-Figueroa, F., Kermit, R., Laborde, C. J., Tanksley, S. D. (1989). Patterns of genetic variation of the genus Capsicum (Solanaceae) in Mexico. Plant Systematics and Evolution, 165(3–4), 159–188. DOI 10.1007/BF00936000. [Google Scholar] [CrossRef]
13. Hernández-Verdugo, S., Guevara-González, R. G., Rivera-Bustamante, R. F., Vázquez-Yanez, C., Oyama, K. (1998). Los parientes silvestres del chile (Capsicum spp.) como recursos genéticos. Boletín de la Sociedad Botánica de México, 62, 171–181. DOI 10.17129/botsci.1559. [Google Scholar] [CrossRef]
14. Hernández-Verdugo, S., Luna-Reyes, R., Oyama, K. (2001). Genetic structure and differentiation of wild and domesticated populations of Capsicum annuum from Mexico. Plant Systematics and Evolution, 226(3–4), 129–142. DOI 10.1007/s006060170061. [Google Scholar] [CrossRef]
15. Hernández-Verdugo, S., González-Rodríguez, A., Sánchez-Peña, P., Casas, A., Oyama, K. (2006). Estructura y diferenciación genética de poblaciones silvestres y domesticadas de chile (Capsicum annuum) del noroeste de México analizada con isoenzimas y RAPDs. Revista Fitotecnia Mexicana, 29(Núm. especial 2), 25–29.
16. Oyama, K., Hernández-Verdugo, S., Sánchez, A. C., González-Rodríguez, A., Sánchez-Peña, P. et al. (2006). Genetic structure of wild and domesticated populations of Capsicum annuum (Solanaceae) from northweste Mexico analized by RAPDs. Genetic Resources and Crop Evolution, 53(3), 553–562. DOI 10.1007/s10722-004-2363-1. [Google Scholar] [CrossRef]
17. Pacheco-Olvera, A., Hernández-Verdugo, S., Rocha-Ramírez, V., Gonzáles-Rodríguez, A., Oyama, K. (2012). Genetic diversity and structure of pepper (Capsicum annuum L.) from Northwestern Mexico analyzed by microsatellite markers. Crop Science, 52(1), 231–241. DOI 10.2135/cropsci2011.06.0319. [Google Scholar] [CrossRef]
18. Dillon, W. R., Golstein, M. (1984). Multivariate analysis: methods and applications. New York USA: Wiley. [Google Scholar]
19. Pigliucci, M., Schlichting, C. D. (1995). Reaction norms of Arabidopsis (Brassivaceace). III. Response to nutrients in 26 populations from worldwide collection. American Journal of Botany, 82(9), 1117–1125. DOI 10.1002/j.1537-2197.1995.tb11582.x. [Google Scholar] [CrossRef]
20. SAS Institute Inc. Cary, North Caroline USA. [Google Scholar]
21. Adebola, P. O., Morakinyo, J. A. (2006). Evaluation of morpho-agronomic variability of wild and cultivated kola (Cola species Schott et Endl.) in South Western Nigeria. Genetic Resources and Crop Evolution, 53(4), 687–694. DOI 10.1007/s10722-004-3558-1. [Google Scholar] [CrossRef]
22. Lewu, F. B., Adebola, P. O., Afolayan, A. J. (2007). Commercial harvesting of Pelargonium sidoides in the Eastern Cape, South Africa: Striking a balance between resource conservation and rural livelihoods. Journal of Arid Environments, 70(2), 380–388. DOI 10.1016/j.jaridenv.2006.12.022. [Google Scholar] [CrossRef]
23. Nooryazdan, H., Serieys, H., Baciliéri, R., David, J., Bervillé, A. (2010). Structure of wild annual sunflower (Helianthus annuus L.) accessions based on agromorphological traits. Genetics Resources and Crop Evolution, 57(1), 27–39. DOI 10.1007/s10722-009-9448-9. [Google Scholar] [CrossRef]
24. Hernández-Verdugo, S., Porras, F., López-España, R. G., Villarreal-Romero, M., Parra-Terraza, S. et al. (2012). Caracterización y variación ecogeográfica de poblaciones de chile silvestre del noroeste de México. Polibotánica, 33, 175–191. [Google Scholar]
25. Orobiyi, A., Loko, L. Y., Sanoussi, F., Agré, A. P., Korie, N. et al. (2018). Agro-morphological characterization of chili pepper landraces (Capsicum annuum L.) cultivated in Northern Benin. Genetic Resources and Crop Evolution, 65(2), 555–569. DOI 10.1007/s10722-017-0553-x. [Google Scholar] [CrossRef]
26. Villota-Cerón, D., Bonilla-Betancourt, M. L., Carmen-Carrillo, H., Jaramillo-Vázquez, J., García Dávila, M. A. (2012). Morphological characterization of Capsicum spp. accesions from the germoplasma collection of Corpoica C.I. Palmira, Colombia. Acta Agronómica, 61(1), 16–26. DOI 10.15446/acag. [Google Scholar] [CrossRef]
27. Hernández-Verdugo, S., López-España, R. G., Sánchez-Peña, P., Villarreal-Romero, M., Parra-Terraza, S. et al. (2008). Variación fenotípica entre y dentro de poblaciones silvestres de chile del noroeste de México. Revista Fitotecnia Mexicana, 31(4), 323–330. [Google Scholar]
28. López-España, R. G., Hernández-Verdugo, S., Parra-Terraza, S., Porras, F., Pacheco-Olvera, A. et al. (2016). Geographical differentiation of wild pepper (Capsicum annuum L. var. glabriusculum) populations from northwestern Mexico. Phyton-International Journal of Experimental Botany, 85, 131–141. [Google Scholar]
29. Pickersgill, B. (1986). Evolution of hierarchical variation patterns under clomestication and their taxonomic treatment. In: Styles, B. T. (ed.Infraespecific classification of wild and cultivated plants, pp. 191–209. Oxford, UK: Oxford University Press. [Google Scholar]
30. Pickersigill, B., Heiser, C. B., McNeil, J. (1979). Numerical taxonomic studies on variation and domestication in some species of Capsicum. In: Hawkes, J. G., Lester, R. N., Skelding, A. D. (eds.Biology and taxonomy of the solanaceae, pp. 679–700. London: Academic Press. [Google Scholar]
31. Palacios-Castro, S., García-Dávila, M. A. (2008). Morphologic characterization of 93 accesions of Capsicum spp of germoplasm bank of National University of Colombia-Palmira. Acta Agronómica, 57(4), 247–252. DOI 10.15446/acag. [Google Scholar] [CrossRef]