
 Submit a Paper
Submit a Paper Propose a Special lssue
Propose a Special lssue Open Access
Open Access
REVIEW
Unlocking the potential of tumor-targeting peptides in precision oncology
1 University Institute of Medical Lab Technology, Faculty of Allied Health Sciences, The University of Lahore, Lahore, 54590, Pakistan
2 Centre for Applied Molecular Biology, 87-West Canal, Bank Road, University of the Punjab, Lahore, 53700, Pakistan
* Corresponding Authors: HAFIZ MUHAMMAD REHMAN. Email: ; HAMID BASHIR. Email:
(This article belongs to the Special Issue: Advances in Cancer Pharmacology)
Oncology Research 2025, 33(7), 1547-1570. https://doi.org/10.32604/or.2025.062197
Received 12 December 2024; Accepted 02 April 2025; Issue published 26 June 2025
 View Full Text
View Full Text Download PDF
Download PDFAbstract
Targeted cancer therapy has emerged as a promising alternative to conventional chemotherapy, which is often plagued by poor selectivity, off-target effects, and drug resistance. Among the various targeting agents in development, peptides stand out for their unique advantages, including minimal immunogenicity, high tissue penetration, and ease of modification. Their small size, specificity, and flexibility allow them to target cancer cells while minimizing damage to healthy tissue selectively. Peptide-based therapies have shown great potential in enhancing the efficacy of drug delivery, improving tumor imaging, and reducing adverse effects. With cancer responsible for millions of deaths worldwide, the development of peptide-based therapeutics offers new hope in addressing the limitations of current treatments. As detailed studies on different aspects of targeting peptides are crucial for optimizing drug development, this review provides a comprehensive overview of the literature on tumor-targeting peptides, including their structure, sources, modes of action, and their application in cancer therapy—both as standalone agents and in fusion drugs. Additionally, various computational tools for peptide-based tumor-targeting drug design and validation are explored. The promising results from these studies highlight peptides as ideal candidates for targeted cancer therapies, offering valuable insights for researchers and accelerating the discovery of novel anti-tumor peptide base drug candidates.Keywords
Cancer remains one of the leading causes of death worldwide, with approximately 20 million new cases and over 9.7 million deaths till 2022 [1]. Despite advancements in early detection and therapeutic interventions, cancer treatment continues to be a significant challenge for clinicians and researchers. Traditional therapies such as chemotherapy, radiation therapy, and surgery are often limited by their lack of selectivity toward tumor cells, leading to severe side effects and damage to healthy tissue [2]. Chemotherapeutic agents, while effective at targeting rapidly dividing cells, often result in off-target toxicity, as they do not discriminate between cancerous and healthy tissues. This lack of specificity frequently leads to adverse effects, including hair loss, nausea, and immune suppression [3]. Furthermore, many cancer cells develop resistance to chemotherapy over time through mechanisms such as drug efflux, gene mutations [4], or alterations in drug metabolism [5]. Targeted cancer therapies have emerged as a promising solution to these challenges. Unlike traditional chemotherapeutics, which non-selectively attack rapidly dividing cells, targeted therapies are designed to interact with specific molecules involved in the growth and survival of cancer cells, sparing healthy tissues and reducing systemic toxicity [6]. These therapies often exploit unique molecular signatures—such as overexpressed receptors, mutated proteins, or altered signaling pathways—that distinguish cancer cells from their normal cells [7]. These therapies offer the potential for improved efficacy and reduced toxicity by selectively attacking cancer cells while minimizing damage to healthy tissues. However, the development of such therapies requires a deep understanding of the molecular mechanisms underlying tumorigenesis and the identification of tumor-specific markers. In recent years, peptides have emerged as highly promising candidates for targeted cancer therapy. These small, versatile molecules can be engineered to selectively bind to receptors that are overexpressed on the surface of tumor cells [8]. Due to their small size, low immunogenicity, and exceptional tissue penetrability, peptides are particularly attractive for drug delivery systems [9]. One of the earliest successes in targeted cancer treatment was imatinib, a tyrosine kinase inhibitor that targets the breakpoint cluster region and Abelson murine leukemia (BCR-ABL) fusion protein in chronic myeloid leukemia (CML) [10]. This shift toward targeted therapies, including peptide-based strategies, reflects an increasing understanding of cancer as a complex and heterogeneous disease. By exploiting the molecular differences between cancerous and normal tissues, targeted therapies offer the potential for more effective, less toxic treatments. This approach is central to the concept of precision medicine, where therapies are tailored to the specific molecular profile of each patient’s tumor, paving the way for more personalized and impactful cancer care.
Monoclonal antibodies are engineered to mimic the immune system’s ability to target specific antigens on cancer cells. They have become a keystone of precision oncology due to their ability to block signaling pathways, induce immune-mediated cytotoxicity, or deliver cytotoxic payloads directly to tumor cells [11]. For example, trastuzumab, a mAb targeting human epidermal growth factor receptor 2 (HER2), has significantly improved outcomes in HER2-positive breast cancer [12]. Recent advancements include the development of bispecific antibodies, which can simultaneously engage two different antigens, enhancing their therapeutic efficacy [13]. Monoclonal antibody fragments, such as Fab (antigen-binding fragment) and scFv (single-chain variable fragment), offer advantages over full-length mAbs, including smaller size, better tissue penetration, and reduced immunogenicity [14]. These fragments are particularly useful in targeting solid tumors and have been employed in imaging and drug delivery systems [15]. For instance, nanobodies, derived from camelid heavy-chain antibodies, have shown promise in targeting hard-to-reach tumor sites due to their small size and high stability [16].
Gene therapy involves the introduction, removal, or alteration of genetic material within a patient’s cells to treat disease. In oncology, this approach can be used to replace mutated genes that cause cancer with healthy copies, inactivate genes that promote cancer growth, or introduce new genes to help the body fight cancer. Recent developments have focused on enhancing delivery methods and ensuring the precise integration of therapeutic genes to maximize efficacy and safety [17]. Cell therapy, particularly chimeric antigen receptor T-cell therapy, has revolutionized cancer treatment by modifying a patient’s T cells to express receptors specific to cancer antigens. These engineered T cells are then expanded and reintroduced into the patient to target and destroy cancer cells. Advancements in this field aim to improve the efficacy of chimeric antigen receptor T-cell therapies against solid tumors and reduce associated toxicities [18].
In this review, we aim to provide a detailed analysis of the latest research on tumor-targeting peptides, including their structural features, sources, and modes of action. We also explore the current and potential applications of peptides in cancer therapy, both as standalone treatments and in combination with small-molecule drugs or other therapeutic agents. Furthermore, we discuss the computational tools and techniques used to design peptide-based cancer therapies, offering examples of how these tools have been successfully implemented in drug development. This study will facilitate the discovery of novel peptides and improve the design of effective, targeted cancer treatments, addressing the ongoing challenges in oncology.
Structure of Targeting Peptides
Peptide bonds bind short chains of amino acids together to form peptides. Their stability and biological function are greatly influenced by their structural characteristics, including length, amino acid sequence, and total charge [19]. The peptide’s general characteristics and interactions with target receptors are determined by its fundamental structure, or the arrangement of its amino acids [20]. This sequence can be altered to improve binding specificity and affinity [21]. Alpha-helices and beta-sheets are examples of secondary structures that affect peptide stability and conformational flexibility, both of which are critical for efficient interaction with tumor targets [22].
α-Helical anticancer peptides (ACPs), such as Magainin II from the African clawed frog [23], are extensively studied for their potent anti-tumor activity at micromolar concentrations, targeting negatively charged tumor cell membranes through electrostatic and hydrophobic interactions. These peptides, with a high net positive charge (+2 to +9 C) due to lysine and arginine residues, disrupt tumor cell membranes and induce necrosis [24]. Magainin II has demonstrated efficacy against bladder cancer and lung cancer cells (IC50 110 µg/mL) while being non-toxic to normal epidermal cells [25]. Other α-helical peptides, such as A12L/A20L and Aurein, isolated from frog secretions, also exhibit significant specificity and anti-tumor activity against various cancers, including glioblastoma [26]. The hydrophobic arc dimensions of these peptides are crucial for their selectivity, enabling them to degrade the plasma and mitochondrial membranes of tumor cells while sparing non-cancerous cells. β-pleated sheet ACPs, primarily derived from plants and animals, are structurally stable due to the presence of two or more disulfide bonds. Although less prevalent than α-helical peptides, they exhibit significant anti-tumor activity with minimal toxicity to normal tissues. Peptides like SVS-1 adopt a β-pleated conformation upon contact with tumor cell surfaces, forming pores and disrupting membranes, with activity against cell lines such as A549 (human lung epithelial cell line), KB (subline of the HeLa cervical cancer), MCF-7 (human breast cancer cell), and MDA-MB-436 (MD Anderson-Metastatic Breast-436 cell line) [27]. LfcinB, derived from cattle lactoferrin [28], and HNP-1, targeting PC-3 prostate cancer cells (IC50 2.2 µM), exemplify the potential of β-pleated ACPs. Despite being less potent than α-helical peptides, their lower toxicity makes them promising candidates for anti-cancer drug development. Random coil ACPs, rich in proline and glycine, lack a defined secondary structure and exhibit moderate anti-tumor activity compared to β-pleated sheet peptides. Examples include PR-39, isolated from neutrophils, which shows antiproliferative effects on tumor cells but also affects normal kidney cells, and its mutant PR-35, which reduces cytotoxicity while maintaining biological activity [29]. Peptides like Pep27anal2, with random coil conformations, penetrate cell membranes and induce apoptosis independently of caspase and cytochrome c [30]. Insect-derived random coil peptides display significant therapeutic potential, and Alloferon has demonstrated antiviral and immunomodulatory effects, enhancing NK cell activity and interferon synthesis. Despite their lower cytotoxicity on healthy tissues compared to α-helical and β-pleated ACPs, random coil peptides hold promise for therapeutic applications. Tertiary structure, the three-dimensional folding of the peptide, is critical for precise receptor binding and cellular uptake [31]. By strategically modifying these structural elements, researchers can improve peptide stability, enhance target selectivity, and optimize therapeutic efficacy, making peptides more effective in specifically targeting and treating cancer cells.
High-throughput screening (HTS) in cancer peptide discovery is a technology-driven approach used to rapidly identify peptides with potential therapeutic or diagnostic applications in oncology.
Phage display technology has significantly advanced in recent years, enhancing its application in anticancer peptide discovery [32]. This technique involves engineering bacteriophages to display peptides or proteins on their surface, facilitating the screening of extensive libraries to identify sequences that specifically interact with cancer-related targets [33]. Through a process called biopanning, peptides that bind to immobilized cancer targets—such as receptors, enzymes, or tumor biomarkers—are enriched via iterative rounds of binding, washing, elution, and amplification [34]. Sequencing the phage DNA reveals the peptide sequences, which are subsequently synthesized and evaluated for therapeutic or diagnostic potential [35]. For instance, phage display has been instrumental in identifying tumor-homing peptides that enhance targeted drug delivery and imaging [36]. Despite challenges such as in vitro bias, target accessibility issues, and the need for peptide optimization [37], phage display continues to be a powerful tool in oncology research, contributing to the discovery of novel peptides for cancer therapy and diagnosis.
Yeast display and combinatorial peptide library
Yeast display technology facilitates the identification of peptides with high specificity and affinity for cancer-related targets. It has been widely used in anticancer peptide discovery due to its robustness, ability to express complex peptides, and compatibility with fluorescence-activated cell sorting (FACS) for precise selection [38]. The system typically involves the fusion of a peptide library to a scaffold protein, such as the Aga2p subunit of the Aga1p–Aga2p adhesion complex in Saccharomyces cerevisiae [39]. This allows peptides to be displayed on the yeast surface while maintaining proper folding and functionality. The displayed peptides are screened against cancer-related targets, such as membrane receptors or tumor-specific antigens, using iterative rounds of selection (biopanning). High-affinity binders are enriched through FACS or magnetic bead-based separation [40]. Unlike phage display, the cost-effectiveness and reproduciblele yeast cells provide post-translational modifications and proper folding, quantitative Screening, library diversity and affinity maturation which enables the generation of large peptide libraries (~109 variants) for iterative selection to improve affinity and specificity [41]. Previously, yeast display technology had identified peptides that inhibit the PD-1/PD-L1 interaction, thereby enhancing T-cell activation against tumors. For instance, researchers have engineered high-affinity variants of the PD-1 ectodomain using yeast surface display. These engineered proteins exhibit superior tumor penetration and efficacy compared to traditional anti–PD-L1 antibodies [42]. However, there are limitations such as size constraints, being more suitable for short peptides and small proteins, and lower display efficiency compared to phage display. Additionally, it may struggle with displaying non-natural peptides containing unnatural amino acids or post-translational modifications [43]. Some other HTS includes peptide nucleic acid (PNA)-encoded solution phase peptide library which offers cost-effective lead ligand optimization and multiple-use potential but is limited by its complex DNA decoding process and non-commercial availability [44]. Peptide microarrays provide cost-effective screening with replicable peptide chips but are restricted in library size and not reusable for subsequent assays [45]. Bacteria-display allows quantitative screening using fluorescence-activated cell sorting (FACS) without requiring reinfection but may suffer from surface interference during peptide binding [46]. Ribosome- or mRNA-display systems enable high library diversity and easy mutagenesis but have low display efficiency [47]. Chemical libraries, such as the one-bead-one-compound approach, facilitate efficient peptide synthesis and screening beyond natural amino acids, but their linker effects remain unpredictable, and they are not suitable for in vivo selection [48]. These methods collectively enhance peptide discovery for biomedical applications, particularly in anticancer research.
Natural peptides derived from various sources have shown significant potential in targeting cancer cells due to their specificity, binding affinity, and therapeutic efficacy. These peptides can be isolated from diverse sources such as animals, plants, microorganisms, and marine organisms, each offering unique properties that can be harnessed for cancer treatment.
The peptides listed in Table 1 demonstrate diverse anti-cancer activities by targeting specific pathways, signalling molecules, or cellular processes across various tumor types. Pardaxin, derived from the Red Sea flatfish, inhibits nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) signaling in murine fibrosarcoma cells, showing significant anti-tumor effects both in vitro and in vivo [49]. AUNP-12, a PD-1 antagonist, enhances immune cell functions and is effective against melanoma, breast, and kidney cancers [50]. Dermaseptin-PD-2, sourced from the Phyllomedusine leaf frog, impedes proliferation in glioblastoma cells [51], while XLAsp-P1 from the South African clawed frog exhibits inhibitory activity against breast cancer cells [52]. Melittin, a peptide from honeybee venom, exhibits potent anticancer activity by causing membrane disruption and promoting cell death in various cancer types [53]. Pentadactylin, identified in the pepper frog, demonstrates cytotoxic effects on murine melanoma cells, though with limited specificity [54]. Marine-derived peptides, such as the oyster peptide and TFD glycoprotein from Pacific cod, induce DNA damage and apoptosis in colon and prostate cancer cells, respectively [55]. Blood clam-derived peptides, along with those from skate, freshwater crocodile [56], and Sinonovacula constricta, show potent cytotoxic and apoptotic activities across a range of cell lines, including cervical, lung, and prostate cancers [57]. Notably, lactoferrin fragments regulate cell death in leukemia and breast cancer cells [58], while A5-1 peptide inhibits integrin-mediated tumor neovascularization [59]. Additionally, Diazonamide A from an unspecified animal source inhibits tubulin polymerization, showing efficacy in breast cancer cells [60]. Collectively, these peptides represent promising candidates for targeted cancer therapies, leveraging mechanisms such as apoptosis, DNA damage, cytotoxicity, and immune modulation.
Plant-derived peptides often exhibit potent anticancer activities by targeting key molecular pathways and inducing specific cellular mechanisms. NDF, isolated from the common bean suppresses colon cancer cell proliferation by modulating proteins involved in cell cycle regulation, such as p53 and p21, and promoting apoptosis through pathways involving BAD, cytC, and caspase-3 [61]. Chickpea-derived CPe-III elevates p53 levels, effectively inhibiting breast cancer cell proliferation [62]. Similarly, RSP-4–3-3 from rapeseed induces apoptosis and demonstrates antiproliferative action in liver cancer cells. P51, derived from plants, binds strongly to HER2 receptors, enhancing the precision of breast cancer therapies [63]. Bitter gourd peptide BG-4 promotes controlled cell death in colon and ovarian cancer [64], while olive seed and walnut residual peptideexhibit cytotoxic effects on prostate and breast cancers through apoptosis and cellular self-digestion [65]. Buckwheat-derived TBWSP31 regulates cell death in breast cancer [66], and the Cycas revoluta peptide induces apoptosis in colon carcinoma and epidermoid cancer cells [67]. Lunasin, a versatile peptide from soy and wheat, inhibits chromatin attachment and prevents malignant transformation in cancer cells triggered by oncogenes or chemical carcinogens [68]. Collectively, these peptides, as listed in Table 2, demonstrate diverse mechanisms such as apoptosis, cell cycle regulation, and receptor-specific targeting, highlighting their potential as natural therapeutic agents for various cancers.
Microbial-derived peptides in Table 3 exhibit a wide range of anti-cancer properties, targeting diverse pathways and tumor types. iPep624, derived from microbial sources, inhibits the transcription factor EN1, effectively targeting breast cancer [79]. Azurin, a bacterial protein, modulates p53 activity to deregulate proliferation and induce apoptosis across cancers like melanoma and breast carcinoma [83]. Similarly, Exotoxin A and Diphtheria toxin inhibit protein synthesis via ADP-ribosylation of elongation factor 2 (EF-2), showing efficacy against cancers such as pancreatic, melanoma, and glioblastoma [84]. Peptides like Pep27anal2 induce apoptosis and permeabilization in multiple cancers [99], while Entap promotes autophagic apoptosis in adenocarcinomas of the gastric, cervix, and mammary gland [86]. Actinomycin D [88] and Bleomycin [89], derived from Actinomyces and Streptomyces species, cause DNA damage through intercalation and free radical generation, targeting cancers such as sarcomas and ovarian carcinomas. Bestatin inhibits aminopeptidase N and functions as an adjuvant in metastasizing lymphoma models [93,94]. Novel peptides like Laterosporulin 10 and Pediocin K2a2-3 demonstrate dual apoptosis and necrosis induction, with potential in colon and breast cancer therapies [91]. Unique compounds like Apratoxin A from Lyngbya majuscula halt the cell cycle in G1 phase [97], while Beauvericin from Fusarium induces caspase-mediated apoptosis [98]. Collectively, these peptides highlight the therapeutic promise of microbial bioactive compounds in oncology.
Synthetic peptides are a diverse class of therapeutic agents designed to target specific cancer-related pathways and structures, offering precision in treatment. M204C4 minimizes pancreatic cancer cell invasion by targeting matrix metalloproteinase-2 (MMP-2) [100]. Prostate-homing peptides deliver pro-apoptotic effects to prostate cancer tissues [101]. PNC-27 induces membrane lysis in breast cancer cells [102], while HRAP inhibits HER2-overexpressing cancer cell proliferation [103]. MAP-04-03 suppresses growth and cell migration in breast cancer, and Lyp-1 selectively binds tumor lymphatics and cells without affecting normal vessels [104]. D-K6L9 disrupts tumor development, and GTI enhances drug delivery specificity to prostate cancer cells by targeting prostate-specific membrane antigen (PSMA) [105]. Peptides like A5G81 promote cell attachment via integrins [106], while LinTT1 targets tumor endothelial cells through mitochondrial protein p32, aiding nanosystem-based therapies [107]. Pep42 targets glucose-regulated protein 78 (GRP78) in melanoma cells offering specificity, while Bld-1 binds selectively to bladder tumor tissues, enabling targeted diagnostics and treatment [108]. ATN-161 and Cilengitide inhibit integrin interactions, reducing tumor cell adhesion and angiogenesis in prostate and various cancers [109]. Anti-angiogenic peptides such as CGNSNPKSC suppress blood vessel growth in gastric cancer [110], while cyclic nonapeptides target aminopeptidase-P to localize conjugated drugs in breast cancer tissue [111]. The binding peptide TLTYTWS reduces angiogenesis in lung cancer models, and TM peptide inhibits tumor growth, angiogenesis, and migration in breast cancer by targeting neuropilin-1 (NRP1) receptors [112]. Together, these peptides exemplify the versatility of synthetic molecules in addressing cancer complexities through tailored mechanisms as listed in Table 4.
Stapled peptides as anti-cancer agent
Stapled peptides are designed by introducing chemical cross-links, or “staples,” to stabilize their α-helical structures, which are crucial for binding to target proteins. This stabilization enhances their resistance to proteolytic degradation and improves cell membrane permeability, making them effective in modulating intracellular protein-protein interactions (PPIs) [113]. The synthesis of stapled peptides commonly involves ring-closing metathesis (RCM) to introduce hydrocarbon staples. This method incorporates non-natural amino acids with olefinic side chains into the peptide sequence, followed by a catalytic reaction to form the staple. Alternative stapling techniques, such as lactam bridges and disulfide bonds, have also been explored to enhance peptide stability and functionality [114]. They have shown significant promise in cancer therapy. For instance, a stapled peptide targeting the MDM2-p53 interaction demonstrated potent antitumor activity by reactivating p53 function in cancer cells [115]. Additionally, stapled peptides designed to inhibit the interaction between the transcription factor STAT3 and its co-activators have been developed, leading to the suppression of tumor growth in preclinical models [116]. The primary advantages of stapled peptides include enhanced stability, improved cell permeability, and the ability to target PPIs with high specificity. However, challenges such as optimizing their pharmacokinetic properties and ensuring efficient delivery to target tissues remain [117]. This class of peptides represent promising anticancer agents, offering the ability to modulate challenging PPIs with enhanced stability and specificity.
Certain tumor-suppressive peptides, such as LL-37 [118] and melittin [119], exhibit amphipathic and cationic properties that enable them to interact with the negatively charged membranes of cancer cells. LL-37, a human cathelicidin peptide, and BMAP-28, a bovine-derived peptide, destabilize these membranes due to their electrostatic interactions, ultimately leading to cell lysis. Cancer cells, with their altered lipid composition and higher negative charge, are more vulnerable to these peptides compared to normal cells (Fig. 1). This selective mechanism allows these peptides to target cancer cells effectively while sparing healthy cells, demonstrating their potential for anticancer therapy.
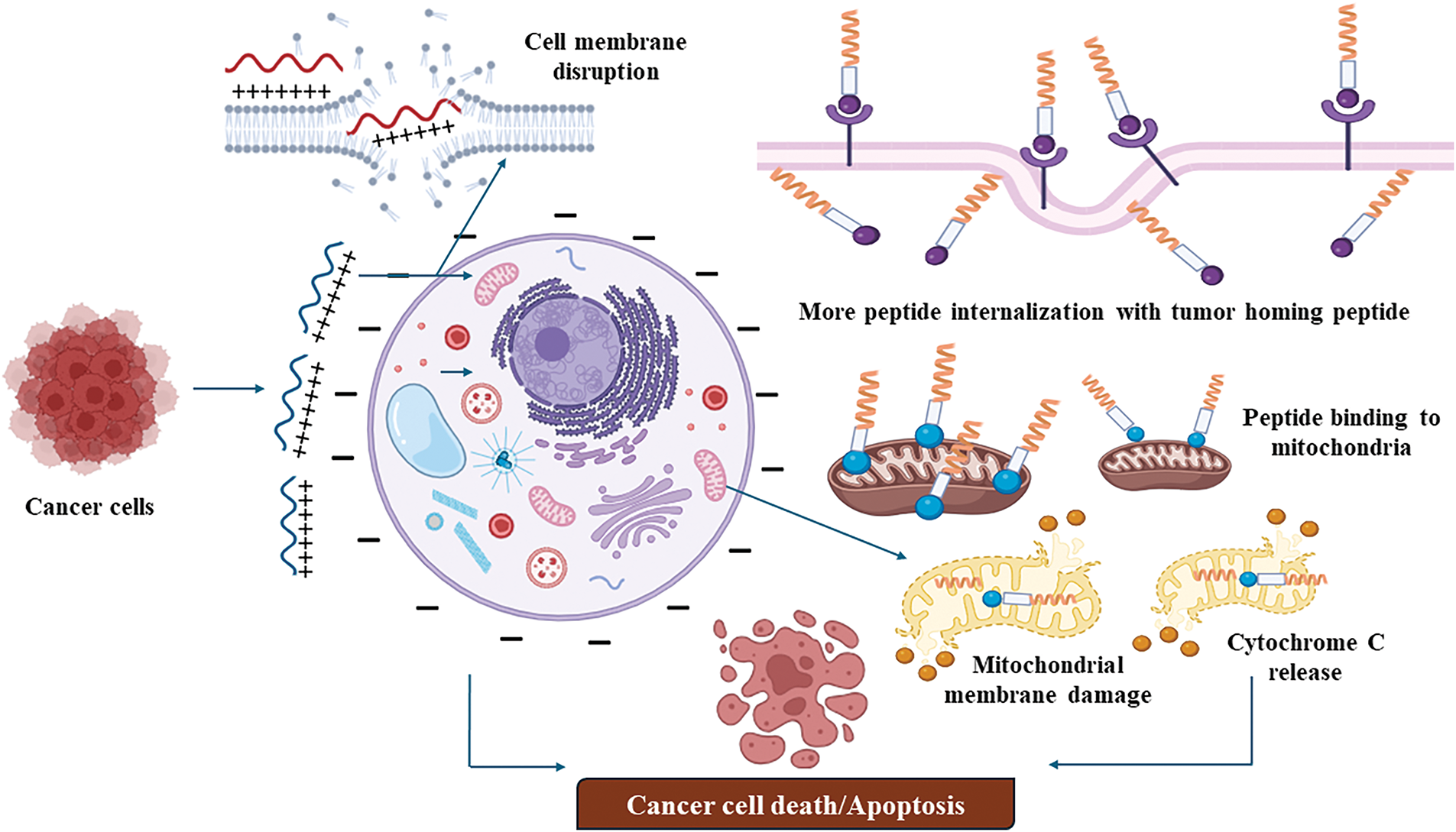
Figure 1: Membrane disrupting mechanism of tumor targeting peptides (created on Biorender, accessible at https://biorender.com/).
Enhanced internalization through tumor-homing peptides
Tumor-homing peptides enhance the targeted delivery and therapeutic efficacy of anticancer peptides by recognizing specific markers or receptors overexpressed on tumor cells or within the tumor microenvironment. These peptides bind selectively to these markers, ensuring precise delivery and deep penetration into tumor tissues, overcoming barriers such as the extracellular matrix. For example, RGD peptides (arginine-glycine-aspartic acid) target integrins, such as αvβ3 and αvβ5, which are commonly overexpressed on the surface of tumor cells and tumor vasculature [120]. Similarly, iRGD peptides, which contain a tumor-penetrating motif, bind to integrins and subsequently interact with neuropilin-1 receptors, facilitating deeper tissue penetration and intracellular delivery of anticancer agents [121]. Another example is F3 peptide, which binds to nucleolin, a marker overexpressed on tumor cells and angiogenic blood vessels, enhancing the uptake of therapeutic agents specifically into cancerous tissues [122] (Fig. 2). These mechanisms ensure selective targeting and efficient therapeutic action with minimal impact on healthy cells.
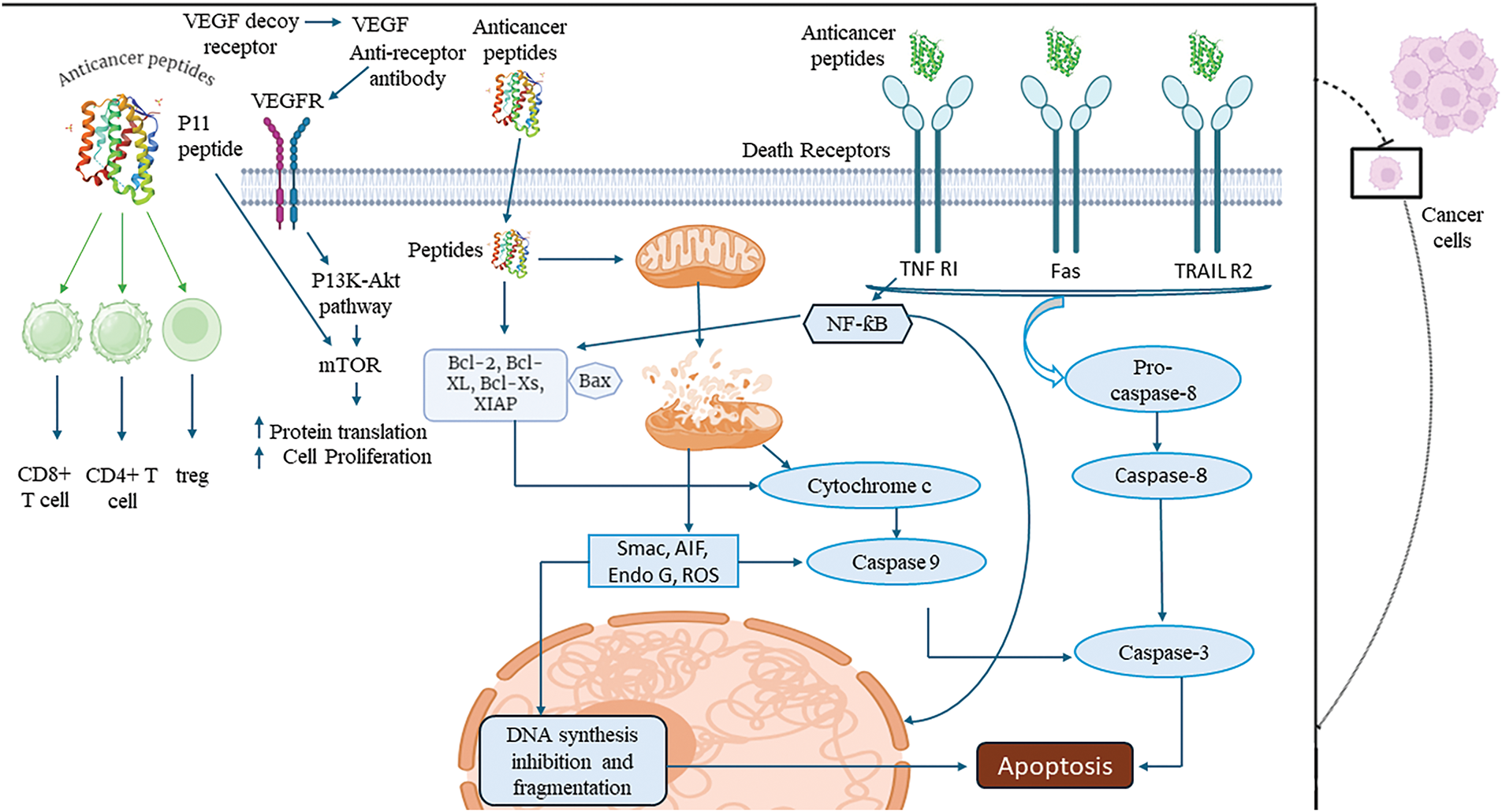
Figure 2: An overview of apoptotic mechanisms of tumor targeting peptides (created on Biorender, accessible at https://biorender.com/). VEGFR: vascular endothelial growth factor receptor, TNF R1: tumor necrosis factor receptor 1, TRAIL R2: tumor necrosis factor-related apoptosis-inducing ligand receptor 2, mTOR: mechanistic target of rapamycin, Bcl-2: B-cell lymphoma 2, Bcl-xL: B-cell lymphoma-extra large, Caspase-8: cysteine-aspartic protease 8, CD8+ T cell: cluster of differentiation 8 positive T cell, CD4+ T cell: cluster of differentiation 4 positive T cell, Treg: regulatory T cell, Smac: second mitochondria-derived activator of caspases, AIF: apoptosis-inducing factor, Endo G: endonuclease G, ROS: reactive oxygen species, Caspase-3: cysteine-aspartic protease 3, DNA: deoxyribonucleic acid, XIAP: X-linked inhibitor of apoptosis protein.
Some peptides specifically target mitochondrial proteins within cancer cells, disrupting mitochondrial membrane integrity and triggering apoptosis. For instance, p53-derived peptides mimic the tumor-suppressor protein p53 and bind to anti-apoptotic proteins like Bcl-2 and Bcl-xL on the mitochondrial membrane, promoting cytochrome C release and intrinsic apoptotic pathway activation [123]. Similarly, KLAKLAK-2, an antimicrobial peptide, targets the mitochondrial membrane directly, compromising its potential and inducing cytochrome C release [124]. Another example is the BID BH3 peptide, which interacts with pro-apoptotic proteins like Bax or Bak, facilitating mitochondrial outer membrane permeabilization (MOMP) and initiating apoptosis [125] (Fig. 2). This strategy has been explored for treating various cancers, including breast cancer, lung cancer, pancreatic cancer, and melanoma, where mitochondrial dysfunction and overexpression of anti-apoptotic proteins are common, ensuring selective and effective cancer cell death.
Apoptosis induction via cytochrome C release
Peptides that disrupt mitochondrial integrity initiate apoptosis by releasing cytochrome C into the cytoplasm, where it binds to apoptotic protease activating factor-1 (Apaf-1) to form the apoptosome, activating caspase-9. This activation triggers caspase-3, leading to DNA fragmentation, chromatin condensation, and programmed cell death. This mechanism selectively eliminates cancer cells while sparing healthy tissues. For example, Bcl-2-converting peptide (Bcl-2CP) binds to anti-apoptotic Bcl-2 family proteins, neutralizing their function and promoting mitochondrial membrane permeabilization [131]. MTP-131 (also known as Bendavia) targets mitochondrial membranes, causing disruption and cytochrome C release, effectively inducing apoptosis [132]. Another example is MAC (Mitochondrial apoptosis-induced channel), which disrupts the mitochondrial membrane potential and triggers cytochrome C release. This strategy has demonstrated potential in treating cancers such as prostate cancer, breast cancer, gastric cancer, and multiple myeloma, where mitochondrial resistance mechanisms are frequently observed [133] (Fig. 2). This strategy has demonstrated potential in treating cancers such as prostate cancer, breast cancer, gastric cancer, and multiple myeloma, where mitochondrial resistance mechanisms are frequently observed.
Certain peptides inhibit the vascular endothelial growth factor (VEGF) signaling pathway by specifically binding to the VEGF receptor (VEGFR), preventing its activation. This inhibition blocks the downstream PI3K-Akt-mTOR signaling cascade, a critical pathway for tumor angiogenesis, growth, and survival [134]. By disrupting angiogenesis, these peptides deprive tumors of essential nutrients and oxygen, effectively curbing tumor growth and reducing metastatic potential. For example, SP5.2, a peptide antagonist of VEGFR-2, binds to the receptor and inhibits VEGF-mediated angiogenic signaling [135]. Similarly, A7R peptide targets VEGFR-2 and neuropilin-1, impairing endothelial cell proliferation and migration necessary for blood vessel formation [136]. CBO-P11 peptide binds directly to VEGFR-1, suppressing VEGF signaling and reducing vascular permeability [137] (Fig. 2). This strategy has shown promise in targeting cancers such as colorectal cancer, breast cancer, non-small cell lung cancer, and glioblastoma, where VEGF-driven angiogenesis plays a pivotal role in tumor progression and metastasis.
Peptides targeting death receptors such as TNF-RI, Fas, and TRAIL R2 on the surface of cancer cells activate the extrinsic apoptotic pathway. These peptides bind to death receptors, inducing receptor trimerization and recruiting adaptor proteins like FADD (Fas-associated death domain). This process facilitates the conversion of procaspase-8 into active caspase-8, which subsequently activates caspase-3. Activated caspase-3 executes apoptosis by cleaving essential structural and regulatory proteins, ultimately leading to cancer cell death [138]. This pathway effectively bypasses some resistance mechanisms that cancer cells employ to evade intrinsic apoptosis. For example, Apo-2L/TRAIL-mimetic peptides mimic the TRAIL ligand and bind specifically to TRAIL R2 (DR5), inducing selective apoptosis in cancer cells without affecting normal cells [139]. Similarly, FasL-derived peptides activate the Fas receptor, promoting apoptotic signaling. Another example is TANDEM peptide, which targets TNF-RI, triggering apoptotic cascades in cancer cells [140] (Fig. 2). This strategy is particularly effective in treating cancers such as colorectal cancer, pancreatic cancer, non-small cell lung cancer, and prostate cancer, where death receptor expression is upregulated, making these tumors susceptible to extrinsic apoptotic activation.
Some tumor-suppressive peptides enhance the activity of immune effector cells, such as CD8+ cytotoxic T cells and D4+ helper T cells, while simultaneously suppressing immunosuppressive regulatory T cells (Tregs). By modulating the tumor microenvironment, these peptides reprogram the immune response, boosting the body’s natural ability to recognize and eliminate cancer cells. This dual action strengthens antitumor immunity and contributes to long-term tumor suppression by promoting immune surveillance. For example, cytotoxic T lymphocyte-associated antigen 4 (CTLA-4)-blocking peptides, such as specific peptide fragments derived from ipilimumab, enhance CD8+ T cell activation by inhibiting CTLA-4-mediated immune suppression [141]. MAGE-A3-derived peptides stimulate CD4+ T helper cells, enhancing cytokine production and promoting cytotoxic T cell responses [142]. Additionally, forkhead box P3 (FOXP3)-targeting peptides reduce Treg activity, reversing the immunosuppressive environment that allows tumor cells to evade immune detection [143] (Fig. 2). This strategy is effective in cancers such as melanoma, non-small cell lung cancer, renal cell carcinoma, and head and neck squamous cell carcinoma, where immune evasion is a significant barrier to effective treatment. By enhancing effector T cell activity and suppressing Tregs, these peptides provide a potent mechanism for boosting the immune system’s antitumor response.
DNA synthesis inhibition and fragmentation
Peptides targeting cancer cell nuclei interfere with DNA synthesis, a critical process for tumor cell proliferation. By localizing to the nucleus, these peptides induce DNA fragmentation, disrupt the replication machinery, and halt cancer cell growth [144]. This mechanism complements other apoptotic pathways, ensuring that cancer cells are eliminated through multiple avenues, thereby enhancing therapeutic efficacy. NLS (nuclear localization signal)-tagged pro-apoptotic peptides specifically target the nucleus to induce DNA fragmentation and inhibit replication enzymes like DNA polymerase [145]. BuGZ peptide derivatives disrupt the assembly of the spindle checkpoint complex, leading to DNA replication stress and cell death [146]. Intranuclear lytic peptides (e.g., LTX-315) penetrate the nucleus, disrupt chromatin structure, and induce DNA damage, leading to apoptosis [147]. Another example is TAT-conjugated DNA-intercalating peptides, which penetrate the nucleus and bind to specific sequences, preventing DNA polymerase activity [148] (Fig. 2). This strategy is effective in cancers such as breast cancer, glioblastoma, leukemia, and cervical cancer, where rapid DNA synthesis drives tumor progression. By halting replication and inducing DNA damage, nuclear-targeting peptides provide a targeted and potent approach to cancer therapy either as standalone treatments or in combination or in combination with other therapeutic agents.
Microenvironment barriers to peptide therapy
The tumor microenvironment (TME) significantly influences the efficacy of peptide-based cancer therapies which can affect both the targeting capabilities and pharmacokinetics of therapeutic peptides. Tumor hypoxia, a condition of reduced oxygen levels, is a hallmark of the TME. Hypoxic conditions can lead to the stabilization of hypoxia-inducible factors, which in turn modulate the expression of various genes involved in angiogenesis, metabolism, and cell survival. These changes can alter the behavior of cancer cells and the surrounding stroma, potentially impacting the distribution and effectiveness of peptide-based therapies [149]. The TME is often characterized by an acidic extracellular pH, resulting from increased glycolysis and subsequent lactate production by cancer cells. This acidic environment can affect the stability and activity of therapeutic peptides. However, it also presents an opportunity for designing pH-responsive delivery systems. For example, pH (low) insertion peptides can exploit the acidic conditions to selectively target tumor cells, enhancing the delivery and efficacy of peptide-based treatments [150]. The extracellular matrix in tumors is often denser and more rigid than in normal tissues, due to increased deposition and cross-linking of matrix components. This dense matrix can act as a physical barrier, hindering the penetration and distribution of therapeutic peptides within the tumor. Additionally, remodeling of dense matrix can influence tumor progression and metastasis, further complicating treatment strategies [151]. All these conditions can pose significant challenges to peptide-based cancer therapies. However, by designing peptides and delivery systems that specifically target these features, it is possible to enhance therapeutic efficacy and overcome some of the barriers.
Advanced peptide delivery in cancer therapy
Advanced peptide drug delivery systems, including nanoparticles, liposomes, and polymers, have been developed to enhance the targeting, stability, and therapeutic efficacy of peptides in cancer treatment.
Nanoparticles, typically ranging from 10 to 100 nm, offer several advantages for peptide delivery [152] including enhanced stability by encapsulating peptides within nanoparticles protecting them from enzymatic degradation and premature clearance, thereby prolonging their circulation time. Another strategy is surface modifications with targeting ligands, which enable nanoparticles to bind specifically to tumor cells, improving the precision of drug delivery. For instance, conjugating nanoparticles with peptides like RGD can facilitate the targeting of integrin receptors overexpressed on tumor cells [153]. Nanoparticles can be engineered to release their payload in response to specific stimuli, such as pH changes or enzymatic activity, ensuring that peptides are released at the tumor site [154]. One example is the targeted delivery of anticancer peptides like melittin by using poly lectic-co-glycolic acid nanoparticles, which protect them from enzymatic degradation and enable controlled release in tumors [155].
Liposomes are spherical vesicles composed of lipid bilayers that can encapsulate peptides and are well-tolerated by the body, reducing the risk of adverse reactions [152]. Liposomes can improve the penetration of peptides into tumor tissues by facilitating cellular uptake and further functionalizing liposomes with targeting peptides or antibodies can enhance specificity to tumor cells, thereby increasing therapeutic efficacy [156]. Doxil, a liposome-based system encapsulating doxorubicin, enhances drug biocompatibility, reduces toxicity, and improves tumor penetration. Similar liposomal approaches protect therapeutic peptides and enable targeted cancer cell delivery [157].
In recent years, natural and synthetic polymer-based delivery systems have emerged to enhance drug targeting, prolong circulation, and enable controlled release. These biodegradable and biocompatible polymers improve drug stability and reduce side effects through mechanisms like adsorption, conjugation, and internal loading [158]. Polymers can be synthesized with various functional groups, allowing for customization of drug delivery systems. Many are biodegradable, minimizing long-term toxicity concerns. Polymers can be designed to respond to specific stimuli, such as pH or temperature, enabling controlled release of peptides at the tumor site [159]. Polymeric micelles are another effective system for peptide drug delivery. For instance, pluronic F-127, a triblock copolymer, has been used to form micelles for encapsulating peptides like melittin for targeted cancer therapy. These micelles are capable of responding to external stimuli such as pH changes in the tumor microenvironment, leading to controlled release of the peptide specifically at the tumor site [160].
Applications in Cancer Therapy: FDA-Approved Anti-Cancer Peptides
This section provides a comprehensive overview of FDA-approved ACPs, highlighting their manufacturers, target tissues, and mechanisms of action. The use of peptides underscore the potential of targeted cancer therapies in modern oncology, offering advantages such as specificity, reduced off-target effects, and improved therapeutic outcomes. Carfilzomib (Kyprolis) produced by Amgen, Carfilzomib is a proteasome inhibitor targeting malignant plasma cells, making it a key treatment for multiple myeloma. By disrupting protein degradation pathways essential for tumor survival, it induces cancer cell apoptosis. Its success underlines the potential of proteasome inhibitors in hematologic malignancies [161]. SomaKit TOC manufactured by SomaKit TOC, this drug is specifically designed for gastroenteropancreatic neuroendocrine tumors. Its precise targeting of somatostatin receptors enhances diagnostic and therapeutic strategies for this rare cancer type, showcasing the utility of receptor-targeted peptides [162]. Venetoclax (Venclexta): AbbVie developed Venetoclax as a selective inhibitor of BCL-2, a protein that prevents apoptosis in cancer cells. By targeting this anti-apoptotic pathway, Venetoclax has proven effective in treating hematologic cancers like chronic lymphocytic leukemia, reflecting the promise of apoptosis-restoring therapies [163]. Osimertinib (Tagrisso): Produced by AstraZeneca, Osimertinib is a third-generation tyrosine kinase inhibitor (TKI) targeting EGFR mutations, a frequent driver of non-small cell lung cancer. It offers benefits over first- and second-generation TKIs, such as improved blood-brain barrier penetration and reduced resistance development, making it a cornerstone in lung cancer therapy [164]. Doxil, a liposomal formulation of doxorubicin by Sequus Pharmaceuticals, Doxil is used for breast cancer, ovarian cancer, and other solid tumors. Encapsulation within liposomes minimizes systemic toxicity and cardiac side effects, exemplifying how nanotechnology enhances peptide-based chemotherapies’ safety profiles [165]. Aprepitant (Emend®): Developed by merck sharp and Dohme BV, Aprepitant is primarily used for its anti-emetic properties in cancer treatment. While it targets neurokinin-1 receptors, its role in lung, neuroendocrine, and gastric cancers demonstrates the potential of dual-function drugs that address cancer symptoms and progression [166]. Cisplatin and Cisplatinum: Manufactured by Cadila Pharmaceuticals, these platinum-based compounds are staples in chemotherapy for bladder, head and neck, lung, ovarian, and testicular cancers. Despite their broad-spectrum activity, their high toxicity levels highlight the ongoing need for safer alternatives or combination therapies to minimize side effects [167].
Many of these agents work by disrupting critical pathways in cancer cells, such as proteasome activity (Carfilzomib) or apoptotic resistance (Venetoclax). Others, like SomaKit TOC, focus on receptor-specific targeting, illustrating how tailored mechanisms enhance therapeutic efficacy. Although these peptides are FDA-approved, challenges remain, including resistance development (e.g., Osimertinib in lung cancer) and systemic toxicities (e.g., Cisplatin). Advanced formulations like Doxil showcase the potential of nanotechnology to address such limitations. The success of these peptides points to the need for continued research into combination therapies, next-generation formulations, and targeted delivery systems. Expanding their application to underexplored cancers could further enhance their impact on global oncology.
Peptide-based in fusion cancer therapy
Advances in anticancer therapies have introduced novel treatments that leverage antibody-drug conjugates (ADCs), radioligands, and peptide-based agents to target specific cancer pathways with improved efficacy and reduced off-target effects. These innovative approaches integrate conjugated peptides and proteins to enhance tumor specificity and therapeutic potential, marking a significant shift in precision oncology. Some notable examples of such therapies, including their conjugated peptide components and clinical outcomes are discussed in this section. Sacituzumab govitecan, an antibody-drug conjugate targeting Trophoblast cell surface antigen 2 (Trop-2), demonstrated remarkable efficacy in treating metastatic triple-negative breast cancer. A phase 3 trial revealed significant improvements in progression-free survival and overall survival compared to standard chemotherapy [168]. Lutetium-177–Dotatate has emerged as a potent treatment for advanced midgut neuroendocrine tumors when a somatostatin analog peptide was conjugated with Leutetium-177. Clinical trials showed markedly extended progression-free survival and higher response rates compared to high-dose octreotide long acting release (LAR). While mild myelosuppression was noted, the absence of renal toxicity emphasized its safety and therapeutic potential [169]. Radioligand therapy with 177Lu-PSMA-617 (Lutetium-177 joined with a prostate-specific membrane antigen (PSMA)-targeting peptide), combined with standard care, significantly improved progression-free survival and overall survival in patients with advanced PSMA-positive metastatic castration-resistant prostate cancer. Though associated with grade 3 or higher adverse events, such as hematologic toxicities, the treatment did not affect the quality of life, solidifying its role in this patient population [169]. Denileukin diftitox, a fusion protein conjugate, combines diphtheria toxin fragments with interleukin-2 and targeting IL-2 receptors, exhibited efficacy in patients with persistent or recurrent cutaneous T-cell lymphoma. A phase 3 study reported a 30% overall response rate, with durable responses. The treatment was associated with flu-like symptoms, vascular leak syndrome, and transient hepatic enzyme elevations but was considered tolerable [170]. Rovalpituzumab tesirine, a DLL3-targeting antibody-drug conjugate, showed encouraging antitumor activity in DLL3-expressing small-cell lung cancer during phase 1 trials. An 18% objective response rate was observed in patients treated at effective doses, with manageable toxicity profiles, supporting its further exploration [171]. Ramucirumab, in combination with FOLFIRI, significantly improved overall survival in metastatic colorectal cancer patients as a second-line treatment. The RAISE trial reported consistent benefits across subgroups with a manageable safety profile, validating its role in this therapeutic setting [172].Ongoing research in fusion protein technology is set to transform the landscape of cancer treatment, offering more effective and personalized therapeutic options.
Computational Tools for Protein Modeling and Analyzing Their Therapeutic Potential
Protein modeling software and online tools
Protein modeling tools and online platforms play a vital role in understanding the structure-function relationship of biomolecules. AlphaFold 2 [173], integrated into platforms like The Human Protein Atlas, has transformed structural biology by employing artificial intelligence to predict protein 3D structures with near-experimental accuracy. It enables researchers to investigate protein mechanisms, interactions, and pathways efficiently. SWISS-MODEL [174], an automated homology modeling server, identifies structural templates from databases and builds reliable protein models based on sequence alignment [175]. This tool simplifies the modeling process for proteins with known homologous structures. PyMOL [176], widely used for visualizing biomolecules, also includes plugins for specialized tasks such as antigenicity prediction, aiding in vaccine and immunotherapy design. MOE [177] (Molecular Operating Environment) integrates molecular modeling and cheminformatics, providing capabilities for calculating molecular weight, protein-ligand docking, and drug design. Another notable tool, trRosetta [178], uses deep learning to predict inter-residue orientations and distances, facilitating de novo protein structure prediction. Together, these tools provide a comprehensive suite for studying, visualizing, and interpreting protein structures, making them indispensable for structural biology, drug discovery, and bioinformatics research.
Protein structure validation tools
Protein structure validation ensures the accuracy and reliability of modeled or experimentally derived 3D structures. PROCHECK [179] is a gold-standard tool that evaluates stereochemical parameters of protein models, providing detailed Ramachandran plots to highlight deviations in torsion angles [180]. These insights are crucial for identifying potential modeling errors. MolProbity [181], another powerful validation tool, offers a wide range of structural checks, including the detection of steric clashes, assessment of rotamer quality, and evaluation of backbone dihedral angles. Its ability to provide atomic-level validation makes it an essential tool for refining protein structures. Additional tools, such as QMEAN [182], provide a quantitative estimate of model quality by comparing the predicted structure to experimental data, while Verify3D evaluates the compatibility of the 3D structure with its amino acid sequence, ensuring the model’s reliability for functional studies [183,184]. These validation methods are essential steps before proceeding with downstream applications such as docking, molecular dynamics simulations, or experimental studies, as they ensure that the protein structure accurately reflects its biological reality.
Protein docking tools are essential for predicting and analyzing peptide-receptor interactions, offering a range of sophisticated algorithms for accurate binding predictions. ClusPro, a widely used docking tool, applies clustering algorithms to identify energetically favorable protein-protein complexes, making it ideal for studying large macromolecular assemblies [185] HADDOCK [186,187] (High Ambiguity Driven DOCKing) goes beyond rigid-body docking by incorporating experimental data such as nuclear magnetic resonance (NMR) and mutagenesis studies, enabling flexible docking of proteins, DNA, and small molecules. Z-Dock [188] specializes in rigid-body docking for protein-protein interactions and uses a scoring system based on shape complementarity and electrostatics to predict optimal orientations. MOE (Molecular operating environment) offers comprehensive docking capabilities, including flexible ligand handling, advanced scoring functions, and customizable workflows, making it highly versatile for drug discovery projects [189]. Geometry-based docking tools like PatchDock [190] detect surface complementarity between interacting molecules, providing a quick and efficient method for docking large complexes. Chimera [191], while primarily a visualization tool, integrates docking capabilities through plugins like AutoDock Vina, allowing researchers to study binding poses and interaction patterns in detail [192]. These tools cater to diverse docking requirements, facilitating the exploration of biomolecular interactions in structural biology, biochemistry, and drug discovery.
Molecular dynamics (MD) simulations offer a dynamic perspective on biomolecular behavior, revealing insights into conformational flexibility, binding mechanisms, and stability under physiological conditions. GROMACS [193], renowned for its high performance, is extensively used for simulating proteins, lipids, and nucleic acids. Its efficient algorithms and advanced trajectory analysis tools make it a preferred choice for large-scale MD studies. NAMD [194] excels in handling complex systems, leveraging parallel computing to simulate large biomolecular assemblies, such as solvated protein-ligand complexes, with high computational efficiency. Desmond, part of the Schrödinger suite, provides an intuitive user interface combined with high-accuracy molecular dynamics simulations, making it suitable for studying detailed protein-ligand dynamics and exploring conformational changes. AMBER [195] specializes in using precise force fields to simulate biomolecular interactions accurately and supports advanced parameterization for custom molecules. These tools are often complemented by enhanced sampling techniques, such as metadynamics and free energy calculations, to explore rare events and binding affinities [196]. Molecular simulation tools are invaluable for understanding the dynamic nature of biomolecular systems, predicting their behavior under different conditions, and validating hypotheses generated from static models or docking studies.
Key contributions of bioinformatics in anti-cancer drug design of fusion proteins
A vast amount of published literature exists on this topic, and while it is not possible to cover all of it due to the scope of this review, a selection of key studies will be highlighted to emphasize their importance in the field. Rehman et al. (2024) employed in silico methods to develop an Azurin-BR2 chimeric protein, showcasing high binding affinity to p53 and potential apoptotic induction in cancer cells [197]. Similarly, Khalid et al. (2024) designed a Leptulipin-p28 fusion protein, targeting VEGFR and Cadherin receptors, which demonstrated stability and interaction efficacy in molecular dynamics simulations [198]. In another study, Rehman et al. (2023) reported the in-silico design of a melittin-IL-24 fusion protein, validated through structure quality indices and simulation studies, indicating promising therapeutic potential [199]. Azurin’s anticancer efficacy was further emphasized by Aslam et al. (2024), combining in silico predictions and in vitro validations to confirm its cytotoxicity against breast cancer cells [200]. Qureshi et al. (2024) explored an IL-24-P20 fusion protein, revealing exceptional stability and receptor interaction capabilities [201]. Fatima et al. (2024) computationally engineered an IL-15-NGR peptide, demonstrating targeted cancer cell homing and stability through advanced docking and simulation techniques [202]. Moreover, Rehman et al. (2024) introduced an IL-24-NBD peptide fusion, highlighting successful receptor interaction and apoptotic potential [203]. Earlier studies, such as Moghadam et al. (2019), focused on DT390-STxB constructs, presenting optimized codon usage and structural stability for efficient translation and anti-tumor activity [204]. Goleij et al. (2019) demonstrated the robust interaction of Herceptin-based fusion proteins with HER2 receptors, proposing their candidacy as novel anticancer agents [205]. In silico analysis offers a safe and efficient approach to studying infectious organisms, minimizing the risk of exposure to hazardous pathogens. Collectively, these studies validate the critical role of computational approaches in designing and optimizing fusion proteins for cancer therapy, paving the way for targeted and effective treatments.
Conclusion and Future Perspective
Future research in peptide-based cancer therapy should focus on optimizing peptide design, enhancing therapeutic efficacy, and addressing current limitations to improve clinical outcomes. Peptide modifications such as cyclization, pegylation, and the incorporation of non-natural amino acids can enhance stability and prolong circulation time. Advanced delivery strategies, including nanoparticles, liposomes, and polymeric micelles, can improve bioavailability, while stimuli-responsive systems can enable controlled release at tumor sites. Combination therapies with immune checkpoint inhibitors or chemotherapy may further enhance treatment efficacy. Addressing challenges such as drug resistance, off-target effects, and immunogenicity is crucial for successful clinical translation. Additionally, integrating AI-driven computational modeling and personalized medicine approaches can help tailor peptide therapies to individual patient profiles. By overcoming these challenges, peptide-based therapies have the potential to become a key component of future cancer treatment strategies.
Acknowledgement: This paper reflects the authors’ analysis and interpretation of the most relevant and recent publications on the topic, providing insights based on the existing body of literature.
Funding Statement: None.
Author Contributions: The authors confirm their contribution to the paper as follows: study conception and design: Hafiz Muhammad Rehman; draft manuscript preparation: Sidra Ahmad; review and editing: Azeem Sarwar; visualization: Hamid Bashir; supervision: Hafiz Muhammad Rehman. All authors reviewed the results and approved the final version of the manuscript.
Availability of Data and Materials: The data and materials employed in this study can be obtained from the corresponding author upon request.
Ethics Approval: Not applicable.
Conflicts of Interest: The authors declare no conflicts of interest to report regarding the present study.
References
1. Bray F, Laversanne M, Sung H, Ferlay J, Siegel RL, Soerjomataram I, et al. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2024;74(3):229–63. doi:10.3322/caac.21834. [Google Scholar] [PubMed] [CrossRef]
2. Anand U, Dey A, Chandel AKS, Sanyal R, Mishra A, Pandey DK, et al. Cancer chemotherapy and beyond: current status, drug candidates, associated risks and progress in targeted therapeutics. Genes Dis. 2023;10(4):1367–401. doi:10.1016/j.gendis.2022.02.007. [Google Scholar] [PubMed] [CrossRef]
3. DeVita VTJr, Rosenberg SA, Lawrence TS. DeVita, Hellman, and Rosenberg’s cancer: short title: Lippincott Williams & Wilkins; 2022 [cited 2025 Feb 10]. Available from: https://dl.mehrsys.ir/pdfbooks/DeVita,%20Hellman,%20and%20Rosenberg_s%20Cancer%20Principles%20&%20Practice%20of%20Oncology%2010th%20Edition(www.myuptodate.com).pdf. [Google Scholar]
4. Kasimu K, Cui W, Wang Y, Li X, Wang H, Yu X, et al. Current advances on single or multi-omics analysis of esophageal cancer. Adv Life Sci. 2024;11(2):296–304. doi:10.62940/als.v11i2.3106. [Google Scholar] [CrossRef]
5. Garg P, Malhotra J, Kulkarni P, Horne D, Salgia R, Singhal SS. Emerging therapeutic strategies to overcome drug resistance in cancer cells. Cancers. 2024;16(13):2478. doi:10.3390/cancers16132478. [Google Scholar] [PubMed] [CrossRef]
6. Mendelsohn J, Baselga J. Status of epidermal growth factor receptor antagonists in the biology and treatment of cancer. J Clin Oncol. 2003;21(14):2787–99. doi:10.1200/JCO.2003.01.504. [Google Scholar] [PubMed] [CrossRef]
7. Tufail M, Hu J-J, Liang J, He C-Y, Wan W-D, Huang Y-Q, et al. Hallmarks of cancer resistance. iScience. 2024;27(6):109979. doi:10.1016/j.isci.2024.109979. [Google Scholar] [PubMed] [CrossRef]
8. Rehman HM, Naz M, Ghous AG, Malik M, Ahmad S, Bashir H. Computational design and evaluation of a novel temporin 1CEa-IL24 fusion protein for anti-tumor potential. Biomed Res Ther. 2025;12(2):7138–52. doi:10.15419/bmrat.v12i2.959. [Google Scholar] [CrossRef]
9. Muttenthaler M, King GF, Adams DJ, Alewood PF. Trends in peptide drug discovery. Nat Rev Drug Discov. 2021;20(4):309–25. doi:10.1038/s41573-020-00135-8. [Google Scholar] [PubMed] [CrossRef]
10. Druker BJ, Tamura S, Buchdunger E, Ohno S, Segal GM, Fanning S, et al. Effects of a selective inhibitor of the Abl tyrosine kinase on the growth of Bcr-Abl positive cells. Nat Med. 1996;2(5):561–6. doi:10.1038/nm0596-561. [Google Scholar] [PubMed] [CrossRef]
11. Aboul-Ella H, Gohar A, Ali AA, Ismail LM, Mahmoud AEE-R, Elkhatib WF, et al. Monoclonal antibodies: from magic bullet to precision weapon. Mol Biomed. 2024;5(1):1–44. doi:10.1186/s43556-024-00210-1. [Google Scholar] [PubMed] [CrossRef]
12. Swain SM, Shastry M, Hamilton E. Targeting HER2-positive breast cancer: advances and future directions. Nat Rev Drug Discov. 2023;22(2):101–26. doi:10.1038/s41573-022-00579-0. [Google Scholar] [PubMed] [CrossRef]
13. Asiri AM, Al Ali A, Abu-Alghayth MH. Understanding the role of genetics in tumour and cancer biology. Advan Life Sci. 2025;12(1):35–48. doi:10.62940/als.v12i1.3334. [Google Scholar] [CrossRef]
14. Gezehagn Kussia G, Tessema TS. The potential of single-chain variable fragment antibody: role in future therapeutic and diagnostic biologics. J Immunol Res. 2024;2024(1):1804038. doi:10.1155/2024/1804038. [Google Scholar] [PubMed] [CrossRef]
15. Nelson AL, Dhimolea E, Reichert JM. Development trends for human monoclonal antibody therapeutics. Nat Rev Drug Discov. 2010;9(10):767–74. doi:10.1038/nrd3229. [Google Scholar] [PubMed] [CrossRef]
16. Alexander E, Leong KW. Discovery of nanobodies: a comprehensive review of their applications and potential over the past five years. J Nanobiotechnol. 2024;22(1):661. doi:10.1186/s12951-024-02900-y. [Google Scholar] [PubMed] [CrossRef]
17. Wang L, Liu G, Zheng L, Long H, Liu Y. A new era of gene and cell therapy for cancer: a narrative review. Ann Transl Med. 2023;11(9):321. doi:10.21037/atm-22-3882. [Google Scholar] [PubMed] [CrossRef]
18. Uscanga-Palomeque AC, Chávez-Escamilla AK, Alvizo-Báez CA, Saavedra-Alonso S, Terrazas-Armendáriz LD, Tamez-Guerra RS, et al. CAR-T cell therapy: from the shop to Cancer therapy. Int J Mol Sci. 2023;24(21):15688. doi:10.3390/ijms242115688. [Google Scholar] [PubMed] [CrossRef]
19. Zhang W, Zhang Y, Wang R, Zhang P, Zhang Y, Randell E, et al. A review: development and application of surface molecularly imprinted polymers toward amino acids, peptides, and proteins. Anal Chim Acta. 2022;1234:340319. doi:10.1016/j.aca.2022.340319. [Google Scholar] [PubMed] [CrossRef]
20. Sharma K, Sharma KK, Sharma A, Jain R. Peptide-based drug discovery: current status and recent advances. Drug Discov Today. 2023;28(2):103464. doi:10.1016/j.drudis.2022.103464. [Google Scholar] [PubMed] [CrossRef]
21. Monti A, Vitagliano L, Caporale A, Ruvo M, Doti N. Targeting protein-protein interfaces with peptides: the contribution of chemical combinatorial peptide library approaches. Int J Mol Sci. 2023;24(9):7842. doi:10.3390/ijms24097842. [Google Scholar] [PubMed] [CrossRef]
22. Li T, Lu X-M, Zhang M-R, Hu K, Li Z. Peptide-based nanomaterials: self-assembly, properties and applications. Bioact Mater. 2022;11(5036):268–82. doi:10.1016/j.bioactmat.2021.09.029. [Google Scholar] [PubMed] [CrossRef]
23. Tolos AM, Moisa C, Dochia M, Popa C, Copolovici L, Copolovici DM. Anticancer potential of antimicrobial peptides: focus on buforins. Polym. 2024;16(6):728. doi:10.3390/polym16060728. [Google Scholar] [PubMed] [CrossRef]
24. de Santana CJC, Pires Júnior OR, Fontes W, Palma MS, Castro MS. Mastoparans: a group of multifunctional α-helical peptides with promising therapeutic properties. Front Mol Biosci. 2022;9:824989. doi:10.3389/fmolb.2022.824989. [Google Scholar] [PubMed] [CrossRef]
25. Lehmann J, Retz M, Sidhu SS, Suttmann H, Sell M, Paulsen F, et al. Antitumor activity of the antimicrobial peptide magainin II against bladder cancer cell lines. Eur Urol. 2006;50(1):141–7. doi:10.1016/j.eururo.2005.12.043. [Google Scholar] [PubMed] [CrossRef]
26. Rozek T, Wegener KL, Bowie JH, Olver IN, Carver JA, Wallace JC, et al. The antibiotic and anticancer active aurein peptides from the Australian Bell Frogs Litoria aurea and Litoria raniformis: the solution structure of aurein 1.2. Eur J Biochem. 2000;267(17):5330–41. doi:10.1046/j.1432-1327.2000.01536.x. [Google Scholar] [PubMed] [CrossRef]
27. Gaspar D, Veiga AS, Castanho MA. From antimicrobial to anticancer peptides. A review. Front Microbiol. 2013;4:294. doi:10.3389/fmicb.2013.00294. [Google Scholar] [PubMed] [CrossRef]
28. Wu J, Zang M, Wang S, Qiao X, Zhao B, Bai J, et al. Lactoferricin, an antimicrobial motif derived from lactoferrin with food preservation potential. Crit Rev Food Sci Nutr. 2024;64(25):9032–44. doi:10.1080/10408398.2023.2207650. [Google Scholar] [PubMed] [CrossRef]
29. Xie M, Liu D, Yang Y. Anti-cancer peptides: classification, mechanism of action, reconstruction and modification. Open Biol. 2020;10(7):200004. doi:10.1098/rsob.200004. [Google Scholar] [PubMed] [CrossRef]
30. Pyla M, Kankipati S, Sumithra B, Mishra PK, Mishra B, Mandal SK, et al. Bacterial proteins and peptides as potential anticancer agents: a novel search for protein-based therapeutics. Curr Med Chem. 2024;32(7):1235–63. doi:10.2174/0109298673253414231127162817. [Google Scholar] [PubMed] [CrossRef]
31. Fisher M. Lehninger principles of biochemistry; by David L. Nelson and Michael M. Cox. Chem Educator. 2001;6(1):69–70. doi:10.1007/s00897000455a. [Google Scholar] [CrossRef]
32. Manivannan AC, Dhandapani R, Velmurugan P, Thangavelu S, Paramasivam R, Ragunathan L, et al. Phage in cancer treatment—biology of therapeutic phage and screening of tumor targeting peptide. Expert Opin Drug Deliv. 2022;19(7):873–82. doi:10.1080/17425247.2022.2094363. [Google Scholar] [PubMed] [CrossRef]
33. Zhang K, Tang Y, Chen Q, Liu Y. The screening of therapeutic peptides for anti-inflammation through phage display technology. Int J Mol Sci. 2022;23(15):8554. doi:10.3390/ijms23158554. [Google Scholar] [PubMed] [CrossRef]
34. Wang J, Tan Y, Ling J, Zhang M, Li L, Liu W, et al. Highly paralleled emulsion droplets for efficient isolation, amplification, and screening of cancer biomarker binding phages. Lab Chip. 2021;21(6):1175–84. doi:10.1039/D0LC01146K. [Google Scholar] [PubMed] [CrossRef]
35. Ning AYW, Chun SCY, Hsum YW, Loo WC, Yin ACY, Quan TY. Phage display screening in breast cancer: from peptide discovery to clinical applications. Life Sci. 2024;357:123077. doi:10.1016/j.lfs.2024.123077. [Google Scholar] [PubMed] [CrossRef]
36. Ayo A, Laakkonen P. Peptide-based strategies for targeted tumor treatment and imaging. Pharmaceutics. 2021;13(4):481. doi:10.3390/pharmaceutics13040481. [Google Scholar] [PubMed] [CrossRef]
37. Hutchings CJ, Sato AK. Phage display technology and its impact in the discovery of novel protein-based drugs. Expert Opin Drug Discov. 2024;19(8):1–29. doi:10.1080/17460441.2024.2367023. [Google Scholar] [PubMed] [CrossRef]
38. Boder ET, Wittrup KD. Yeast surface display for screening combinatorial polypeptide libraries. Nat Biotechnol. 1997;15(6):553–7. doi:10.1038/nbt0697-553. [Google Scholar] [PubMed] [CrossRef]
39. Li Y, Wang X, Zhou N-Y, Ding J. Yeast surface display technology: mechanisms, applications, and perspectives. Biotechnol Adv. 2024;76:108422. doi:10.1016/j.biotechadv.2024.108422. [Google Scholar] [PubMed] [CrossRef]
40. Boder ET, Raeeszadeh-Sarmazdeh M, Price JV. Engineering antibodies by yeast display. Arch Biochem Biophys. 2012;526(2):99–106. doi:10.1016/j.abb.2012.03.009. [Google Scholar] [PubMed] [CrossRef]
41. Cherf GM, Cochran JR. Applications of yeast surface display for protein engineering. Yeast Surface Display: Methods Protoc Appl. 2015;1319:155–75. doi:10.1007/978-1-4939-2748-7. [Google Scholar] [CrossRef]
42. Maute RL, Gordon SR, Mayer AT, McCracken MN, Natarajan A, Ring NG, et al. Engineering high-affinity PD-1 variants for optimized immunotherapy and immuno-PET imaging. Proc Natl Acad Sci. 2015;112(47):E6506–14. doi:10.1073/pnas.1519623112. [Google Scholar] [PubMed] [CrossRef]
43. Teymennet-Ramírez KV, Martínez-Morales F, Trejo-Hernández MR. Yeast surface display system: strategies for improvement and biotechnological applications. Front Bioeng Biotechnol. 2022;9:794742. doi:10.3389/fbioe.2021.794742. [Google Scholar] [PubMed] [CrossRef]
44. Kunig VB, Potowski M, Klika Škopić M, Brunschweiger A. Scanning protein surfaces with DNA-encoded libraries. ChemMedChem. 2021;16(7):1048–62. doi:10.1002/cmdc.202000869. [Google Scholar] [PubMed] [CrossRef]
45. Okasha H. Fundamental uses of peptides as a new model in both treatment and diagnosis. Recent Pat Biotechnol. 2024;18(2):110–27. doi:10.2174/1872208317666230512143508. [Google Scholar] [PubMed] [CrossRef]
46. Liu R, Li X, Xiao W, Lam KS. Tumor-targeting peptides from combinatorial libraries. Adv Drug Deliv Rev. 2017;110:13–37. doi:10.1016/j.addr.2016.05.009. [Google Scholar] [PubMed] [CrossRef]
47. Brango-Vanegas J, Leite ML, de Oliveira KBS, da Cunha NB, Franco OL. From exploring cancer and virus targets to discovering active peptides through mRNA display. Pharmacol Ther. 2023;252(2):108559. doi:10.1016/j.pharmthera.2023.108559. [Google Scholar] [PubMed] [CrossRef]
48. Karami Fath M, Babakhaniyan K, Zokaei M, Yaghoubian A, Akbari S, Khorsandi M, et al. Anti-cancer peptide-based therapeutic strategies in solid tumors. Cell Mol Biol Lett. 2022;27(1):33. doi:10.1186/s11658-022-00332-w. [Google Scholar] [PubMed] [CrossRef]
49. Wu S-P, Huang T-C, Lin C-C, Hui C-F, Lin C-H, Chen J-Y. Pardaxin, a fish antimicrobial peptide, exhibits antitumor activity toward murine fibrosarcoma in vitro and in vivo. Mar Drugs. 2012;10(8):1852–72. doi:10.3390/md10081852. [Google Scholar] [PubMed] [CrossRef]
50. Sasikumar PG, Satyam LK, Shrimali RK, Subbarao K, Ramachandra R, Vadlamani S, et al. Demonstration of anti-tumor efficacy in multiple preclinical cancer models using a novel peptide inhibitor (Aurigene-012) of the PD1 signaling pathway. Cancer Res. 2012;72(8_Supplement):2850. doi:10.1158/1538-7445.AM2012-2850. [Google Scholar] [CrossRef]
51. Shi D, Hou X, Wang L, Gao Y, Wu D, Xi X, et al. Two novel dermaseptin-like antimicrobial peptides with anticancer activities from the skin secretion of Pachymedusa dacnicolor. Toxins. 2016;8(5):144. doi:10.3390/toxins8050144. [Google Scholar] [PubMed] [CrossRef]
52. Li S, Hao L, Bao W, Zhang P, Su D, Cheng Y, et al. A novel short anionic antibacterial peptide isolated from the skin of Xenopus laevis with broad antibacterial activity and inhibitory activity against breast cancer cell. Arch Microbiol. 2016;198(5):473–82. doi:10.1007/s00203-016-1206-8. [Google Scholar] [PubMed] [CrossRef]
53. Zhou J, Wan C, Cheng J, Huang H, Lovell JF, Jin H. Delivery strategies for melittin-based cancer therapy. ACS Appl Mater Interfaces. 2021;13(15):17158–73. doi:10.1021/acsami.1c03640. [Google Scholar] [PubMed] [CrossRef]
54. Libério MS, Joanitti GA, Azevedo RB, Cilli EM, Zanotta LC, Nascimento AC, et al. Anti-proliferative and cytotoxic activity of pentadactylin isolated from Leptodactylus labyrinthicus on melanoma cells. Amino Acids. 2011;40(1):51–9. doi:10.1007/s00726-009-0384-y. [Google Scholar] [PubMed] [CrossRef]
55. Guha P, Kaptan E, Bandyopadhyaya G, Kaczanowska S, Davila E, Thompson K, et al. Cod glycopeptide with picomolar affinity to galectin-3 suppresses T-cell apoptosis and prostate cancer metastasis. Proc Natl Acad Sci. 2013;110(13):5052–7. doi:10.1073/pnas.1202653110. [Google Scholar] [PubMed] [CrossRef]
56. Theansungnoen T, Maijaroen S, Jangpromma N, Yaraksa N, Daduang S, Temsiripong T, et al. Cationic antimicrobial peptides derived from Crocodylus siamensis leukocyte extract, revealing anticancer activity and apoptotic induction on human cervical cancer cells. Protein J. 2016;35(3):202–11. doi:10.1007/s10930-016-9662-1. [Google Scholar] [PubMed] [CrossRef]
57. Pan X, Zhao Y-Q, Hu F-Y, Chi C-F, Wang B. Anticancer activity of a hexapeptide from skate (Raja porosa) cartilage protein hydrolysate in HeLa cells. Mar Drugs. 2016;14(8):153. doi:10.3390/md14080153. [Google Scholar] [PubMed] [CrossRef]
58. Arias M, Hilchie AL, Haney EF, Bolscher JG, Hyndman ME, Hancock RE, et al. Anticancer activities of bovine and human lactoferricin-derived peptides. Biochem Cell Biol. 2017;95(1):91–8. doi:10.1139/bcb-2016-0175. [Google Scholar] [PubMed] [CrossRef]
59. Zhou J, Li Y, Huang W, Shi W, Qian H. Source and exploration of the peptides used to construct peptide-drug conjugates. Eur J Med Chem. 2021;224(3):113712. doi:10.1016/j.ejmech.2021.113712. [Google Scholar] [PubMed] [CrossRef]
60. Zhang J-Y, Fu L-W. Advance of several types of important marine antitumor drugs. Yao Xue Xue Bao = Acta Pharmaceutica Sinica. 2008;43(5):435–42. [Google Scholar] [PubMed]
61. Vital DAL, De Mejía EG, Dia VP, Loarca-Piña G. Peptides in common bean fractions inhibit human colorectal cancer cells. Food Chem. 2014;157:347–55. doi:10.1016/j.foodchem.2014.02.050. [Google Scholar] [PubMed] [CrossRef]
62. Xue Z, Wen H, Zhai L, Yu Y, Li Y, Yu W, et al. Antioxidant activity and anti-proliferative effect of a bioactive peptide from chickpea (Cicer arietinum L.). Food Res Int. 2015;77(2):75–81. doi:10.1016/j.foodres.2015.09.027. [Google Scholar] [CrossRef]
63. Wang L, Zhang J, Yuan Q, Xie H, Shi J, Ju X. Separation and purification of an anti-tumor peptide from rapeseed (Brassica campestris L.) and the effect on cell apoptosis. Food Funct. 2016;7(5):2239–48. doi:10.1039/C6FO00042H. [Google Scholar] [PubMed] [CrossRef]
64. Saha M, Chakraborty A, Chatterjee S. Bitter gourd seed: a natural plant unit with immense anti-proliferative activities and active constituents. In: Seeds: anti-proliferative storehouse for bioactive secondary metabolites. Springer; 2024. p. 285–307. doi:10.1007/978-981-97-3014-8_9 [Google Scholar] [CrossRef]
65. Vásquez-Villanueva R, Muñoz-Moreno L, Carmena MJ, Marina ML, García MC. In vitro antitumor and hypotensive activity of peptides from olive seeds. J Funct Foods. 2018;42:177–84. doi:10.1016/j.jff.2017.12.062. [Google Scholar] [CrossRef]
66. Guo X, Zhu K, Zhang H, Yao H. Anti-tumor activity of a novel protein obtained from tartary buckwheat. Int J Mol Sci. 2010;11(12):5201–11. doi:10.3390/ijms11125201. [Google Scholar] [PubMed] [CrossRef]
67. Trinidad-Calderón PA, Varela-Chinchilla CD, García-Lara S. Natural peptides inducing cancer cell death: mechanisms and properties of specific candidates for cancer therapeutics. Molecules. 2021;26(24):7453. doi:10.3390/molecules26247453. [Google Scholar] [PubMed] [CrossRef]
68. Alves de Souza SM, Hernández-Ledesma B, de Souza TLF. Lunasin as a promising plant-derived peptide for cancer therapy. Int J Mol Sci. 2022;23(17):9548. doi:10.3390/ijms23179548. [Google Scholar] [PubMed] [CrossRef]
69. Umayaparvathi S, Meenakshi S, Vimalraj V, Arumugam M, Sivagami G, Balasubramanian T. Antioxidant activity and anticancer effect of bioactive peptide from enzymatic hydrolysate of oyster (Saccostrea cucullata). Biomed Prev Nutr. 2014;4(3):343–53. doi:10.1016/j.bionut.2014.04.006. [Google Scholar] [CrossRef]
70. Yousr MN, Aloqbi AA, Omar UM, Howell NK. Antiproliferative activity of egg yolk peptides in human colon cancer cells. Nutr Cancer. 2017;69(4):674–81. doi:10.1080/01635581.2017.1295087. [Google Scholar] [PubMed] [CrossRef]
71. Zhou J, Zhao M, Tang Y, Wang J, Wei C, Gu F, et al. The milk-derived fusion peptide, ACFP, suppresses the growth of primary human ovarian cancer cells by regulating apoptotic gene expression and signaling pathways. BMC Cancer. 2016;16(1):1–14. doi:10.1186/s12885-016-2281-6. [Google Scholar] [PubMed] [CrossRef]
72. Kim E-Y, Bang JY, Chang S-I, Kang I-C. A novel integrin α5β1 antagonistic peptide, A5-1, screened by protein chip system as a potent angiogenesis inhibitor. Biochem Biophys Res Commun. 2008;377(4):1288–93. doi:10.1016/j.bbrc.2008.10.166. [Google Scholar] [PubMed] [CrossRef]
73. Huang F, Ding G, Yang Z, Yu F. Two novel peptides derived from Sinonovacula constricta inhibit the proliferation and induce apoptosis of human prostate cancer cells. Mol Med Rep. 2017;16(5):6697–707. doi:10.3892/mmr.2017.7418. [Google Scholar] [PubMed] [CrossRef]
74. Castro G, Maria DA, Bouhallab S, Sgarbieri V. In vitro impact of a whey protein isolate (WPI) and collagen hydrolysates (CHs) on B16F10 melanoma cells proliferation. J Dermatol Sci. 2009;56(1):51–7. doi:10.1016/j.jdermsci.2009.06.016. [Google Scholar] [PubMed] [CrossRef]
75. Maraming P, Maijaroen S, Klaynongsruang S, Boonsiri P, Daduang S, Chung J-G, et al. Antitumor ability of KT2 peptide derived from leukocyte peptide of crocodile against human HCT116 colon cancer xenografts. In Vivo. 2018;32(5):1137–44. doi:10.21873/invivo.11356. [Google Scholar] [PubMed] [CrossRef]
76. Geng L, Wang Z, Jia X, Han Q, Xiang Z, Li D, et al. HER2 targeting peptides screening and applications in tumor imaging and drug delivery. Theranostics. 2016;6(8):1261–73. doi:10.7150/thno.14302. [Google Scholar] [PubMed] [CrossRef]
77. Ma S, Huang D, Zhai M, Yang L, Peng S, Chen C, et al. Isolation of a novel bio-peptide from walnut residual protein inducing apoptosis and autophagy on cancer cells. BMC Complement Altern Med. 2015;15(1):1–14. doi:10.1186/s12906-015-0940-9. [Google Scholar] [PubMed] [CrossRef]
78. Bloom AD. BG-4, a bioactive peptide from Momordica charantia, promotes apoptosis in ovarian cancer cells; 2018 [cited 2025 Feb 10]. Available from: https://trace.tennessee.edu/cgi/viewcontent.cgi?article=3196&context=utk_chanhonoproj. [Google Scholar]
79. Beltran A, Graves L, Blancafort P. Novel role of Engrailed 1 as a prosurvival transcription factor in basal-like breast cancer and engineering of interference peptides block its oncogenic function. Oncogene. 2014;33(39):4767–77. doi:10.1038/onc.2013.422. [Google Scholar] [PubMed] [CrossRef]
80. Ueda H, Yamazaki M. Induction of tumor necrosis factor-α in solid tumor region by the orally administered synthetic muramyl dipeptide analogue, romurtide. Int Immunopharmacol. 2001;1(1):97–104. doi:10.1016/S1567-5769(00)00017-5. [Google Scholar] [PubMed] [CrossRef]
81. Zhang HL, Hua HM, Pei YH, Yao XS. Three new cytotoxic cyclic acylpeptides from marine Bacillus sp. Chem Pharm Bull. 2004;52(8):1029–30. doi:10.1248/cpb.52.1029. [Google Scholar] [PubMed] [CrossRef]
82. Kalinovskaya NI, Romanenko LA, Kalinovsky AI, Dmitrenok PS, Dyshlovoy SA. A new antimicrobial and anticancer peptide producing by the marine deep sediment strain “Paenibacillus profundus” sp. nov. Sl 79. Nat Prod Commun. 2013;8(3):1934578X1300800326. doi:10.1177/1934578X1300800326. [Google Scholar] [CrossRef]
83. Goto M, Yamada T, Kimbara K, Horner J, Newcomb M, Gupta TD, et al. Induction of apoptosis in macrophages by Pseudomonas aeruginosa azurin: tumour-suppressor protein p53 and reactive oxygen species, but not redox activity, as critical elements in cytotoxicity. Mol Microbiol. 2003;47(2):549–59. doi:10.1046/j.1365-2958.2003.03317.x. [Google Scholar] [PubMed] [CrossRef]
84. Karpiaski TM, Szkaradkiewicz AK. Anti-cancer peptides from bacteria. Bangladesh J Pharmacol. 2013;8(3):343–8. doi:10.3329/bjp.v8i3.15704. [Google Scholar] [CrossRef]
85. Matsuo Y, Kanoh K, Imagawa H, Adachi K, Nishizawa M, Shizuri Y. Urukthapelstatin A, a novel cytotoxic substance from marine-derived Mechercharimyces asporophorigenens YM11-542. J Antibiot. 2007;60(4):256–60. doi:10.1038/ja.2007.31. [Google Scholar] [PubMed] [CrossRef]
86. Karpiński T. New peptide (Entap) with anti-proliferative activity produced by bacteria of Enterococcus genus. Poland: Poznań University of Medical Sciences Poznań; 2012. [Google Scholar]
87. Murphy JR. Mechanism of diphtheria toxin catalytic domain delivery to the eukaryotic cell cytosol and the cellular factors that directly participate in the process. Toxins. 2011;3(3):294–308. doi:10.3390/toxins3030294. [Google Scholar] [PubMed] [CrossRef]
88. Koba M, Konopa J. Aktynomycyna D i mechanizmy jej działania Actinomycin D and its mechanisms of action. Postepy Hig Med Dosw (Online). 2005;59:290–8. [Google Scholar] [PubMed]
89. Segerman ZJ, Roy B, Hecht SM. Characterization of bleomycin-mediated cleavage of a hairpin DNA library. Biochem. 2013;52(31):5315–27. doi:10.1021/bi400779r. [Google Scholar] [PubMed] [CrossRef]
90. Paiva AD, de Oliveira MD, de Paula SO, Baracat-Pereira MC, Breukink E, Mantovani HC. Toxicity of bovicin HC5 against mammalian cell lines and the role of cholesterol in bacteriocin activity. Microbiol. 2012;158(11):2851–8. doi:10.1099/mic.0.062190-0. [Google Scholar] [PubMed] [CrossRef]
91. Baindara P, Gautam A, Raghava G, Korpole S. Anticancer properties of a defensin like class IId bacteriocin Laterosporulin10. Sci Rep. 2017;7(1):46541. doi:10.1038/srep46541. [Google Scholar] [PubMed] [CrossRef]
92. Villarante KI, Elegado FB, Iwatani S, Zendo T, Sonomoto K, de Guzman EE. Purification, characterization and in vitro cytotoxicity of the bacteriocin from Pediococcus acidilactici K2a2-3 against human colon adenocarcinoma (HT29) and human cervical carcinoma (HeLa) cells. World J Microbiol Biotechnol. 2011;27(4):975–80. doi:10.1007/s11274-010-0541-1. [Google Scholar] [CrossRef]
93. Schorlemmer HU, Bosslet K, Sedlacek HH. Ability of the immunomodulating dipeptide bestatin to activate cytotoxic mononuclear phagocytes. Cancer Res. 1983;43(9):4148–53. [Google Scholar] [PubMed]
94. Aozuka Y, Koizumi K, Saitoh Y, Ueda Y, Sakurai H, Saiki I. Anti-tumor angiogenesis effect of aminopeptidase inhibitor bestatin against B16-BL6 melanoma cells orthotopically implanted into syngeneic mice. Cancer Lett. 2004;216(1):35–42. doi:10.1016/j.canlet.2004.06.050. [Google Scholar] [PubMed] [CrossRef]
95. Wang Z, Zhang X. Isolation and identification of anti-proliferative peptides from Spirulina platensis using three-step hydrolysis. J Sci Food Agric. 2017;97(3):918–22. doi:10.1002/jsfa.7815. [Google Scholar] [PubMed] [CrossRef]
96. Wang X, Zhang X. Separation, antitumor activities, and encapsulation of polypeptide from Chlorella pyrenoidosa. Biotechnol Prog. 2013;29(3):681–7. doi:10.1002/btpr.1725. [Google Scholar] [CrossRef]
97. Kang HK, Choi M-C, Seo CH, Park Y. Therapeutic properties and biological benefits of marine-derived anticancer peptides. Int J Mol Sci. 2018;19(3):919. doi:10.3390/ijms19030919. [Google Scholar] [PubMed] [CrossRef]
98. Tao Y-W, Lin Y-C, She Z-G, Lin M-T, Chen P-X, Zhang J-Y. Anticancer activity and mechanism investigation of beauvericin isolated from secondary metabolites of the mangrove endophytic fungi. Anti-Cancer Agents Med Chem. 2015;15(2):258–66. doi:10.2174/1871520614666140825112255. [Google Scholar] [PubMed] [CrossRef]
99. Lee DG, Hahm K-S, Park Y, Kim H-Y, Lee W, Lim S-C, et al. Functional and structural characteristics of anticancer peptide Pep27 analogues. Cancer Cell Int. 2005;5(1):1–14. doi:10.1186/1475-2867-5-21. [Google Scholar] [PubMed] [CrossRef]
100. Lu G, Zheng M, Zhu Y, Sha M, Wu Y, Han X. Selection of peptide inhibitor to matrix metalloproteinase-2 using phage display and its effects on pancreatic cancer cell lines PANC-1 and CFPAC-1. Int J Biol Sci. 2012;8(5):650–62. doi:10.7150/ijbs.3897. [Google Scholar] [PubMed] [CrossRef]
101. Arap W, Haedicke W, Bernasconi M, Kain R, Rajotte D, Krajewski S, et al. Targeting the prostate for destruction through a vascular address. Proc Natl Acad Sci. 2002;99(3):1527–31. doi:10.1073/pnas.241655998. [Google Scholar] [PubMed] [CrossRef]
102. Sookraj KA, Bowne WB, Adler V, Sarafraz-Yazdi E, Michl J, Pincus MR. The anti-cancer peptide, PNC-27, induces tumor cell lysis as the intact peptide. Cancer Chemother Pharmacol. 2010;66(2):325–31. doi:10.1007/s00280-009-1166-7. [Google Scholar] [PubMed] [CrossRef]
103. Nakajima H, Mizuta N, Sakaguchi K, Fujiwara I, Yoshimori A, Takahashi S, et al. Development of HER2-antagonistic peptides as novel anti-breast cancer drugs by in silico methods. Breast Cancer. 2008;15(1):65–72. doi:10.1007/s12282-007-0018-8. [Google Scholar] [PubMed] [CrossRef]
104. Laakkonen P, Porkka K, Hoffman JA, Ruoslahti E. A tumor-homing peptide with a targeting specificity related to lymphatic vessels. Nat Med. 2002;8(7):751–5. doi:10.1038/nm720. [Google Scholar] [PubMed] [CrossRef]
105. Papo N, Seger D, Makovitzki A, Kalchenko V, Eshhar Z, Degani H, et al. Inhibition of tumor growth and elimination of multiple metastases in human prostate and breast xenografts by systemic inoculation of a host defense-like lytic peptide. Cancer Res. 2006;66(10):5371–8. doi:10.1158/0008-5472.CAN-05-4569. [Google Scholar] [PubMed] [CrossRef]
106. Katagiri F, Ishikawa M, Yamada Y, Hozumi K, Kikkawa Y, Nomizu M. Screening of integrin-binding peptides from the laminin α4 and α5 chain G domain peptide library. Arch Biochem Biophys. 2012;521(1–2):32–42. doi:10.1016/j.abb.2012.02.017. [Google Scholar] [PubMed] [CrossRef]
107. Sharma S, Kotamraju VR, Molder T, Tobi A, Teesalu T, Ruoslahti E. Tumor-penetrating nanosystem strongly suppresses breast tumor growth. Nano Lett. 2017;17(3):1356–64. doi:10.1021/acs.nanolett.6b03815. [Google Scholar] [PubMed] [CrossRef]
108. Lee S-M, Lee E-J, Hong H-Y, Kwon M-K, Kwon T-H, Choi J-Y, et al. Targeting bladder tumor cells in vivo and in the urine with a peptide identified by phage display. Mol Cancer. 2007;5(1):11–9. doi:10.1158/1541-7786.MCR-06-0069. [Google Scholar] [PubMed] [CrossRef]
109. Livant DL, Brabec RK, Pienta KJ, Allen DL, Kurachi K, Markwart S, et al. Anti-invasive, antitumorigenic, and antimetastatic activities of the PHSCN sequence in prostate carcinoma. Cancer Res. 2000;60(2):309–20. [Google Scholar] [PubMed]
110. Hui X, Han Y, Liang S, Liu Z, Liu J, Hong L, et al. Specific targeting of the vasculature of gastric cancer by a new tumor-homing peptide CGNSNPKSC. J Controlled Release. 2008;131(2):86–93. doi:10.1016/j.jconrel.2008.07.024. [Google Scholar] [PubMed] [CrossRef]
111. Shoari A, Khodabakhsh F, Cohan RA, Salimian M, Karami E. Anti-angiogenic peptides application in cancer therapy; a review. Res Pharm Sci. 2021;16(6):559–74. doi:10.4103/1735-5362.327503. [Google Scholar] [PubMed] [CrossRef]
112. Nasarre C, Roth M, Jacob L, Roth L, Koncina E, Thien A, et al. Peptide-based interference of the transmembrane domain of neuropilin-1 inhibits glioma growth in vivo. Oncogene. 2010;29(16):2381–92. doi:10.1038/onc.2010.9. [Google Scholar] [PubMed] [CrossRef]
113. Vadevoo SMP, Gurung S, Lee H-S, Gunassekaran GR, Lee S-M, Yoon J-W, et al. Peptides as multifunctional players in cancer therapy. Exp Mol Med. 2023;55(6):1099–109. doi:10.1038/s12276-023-01016-x. [Google Scholar] [PubMed] [CrossRef]
114. Chandramohan A, Josien H, Yuen TY, Duggal R, Spiegelberg D, Yan L, et al. Design-rules for stapled peptides with in vivo activity and their application to Mdm2/X antagonists. Nat Commun. 2024;15(1):489. doi:10.1038/s41467-023-43346-4. [Google Scholar] [PubMed] [CrossRef]
115. Chang YS, Graves B, Guerlavais V, Tovar C, Packman K, To K-H, et al. Stapled α−helical peptide drug development: a potent dual inhibitor of MDM2 and MDMX for p53-dependent cancer therapy. Proc Natl Acad Sci. 2013;110(36):E3445–54. doi:10.1073/pnas.1303002110. [Google Scholar] [PubMed] [CrossRef]
116. Borghouts C, Delis N, Brill B, Weiss A, Mack L, Lucks P, et al. A membrane penetrating peptide aptamer inhibits STAT3 function and suppresses the growth of STAT3 addicted tumor cells. JAK-STAT. 2012;1(1):44–55. doi:10.4161/jkst.18947. [Google Scholar] [PubMed] [CrossRef]
117. Zhang Q, Wang Z, Mei X, Chen Q, Zhang C. Stapled peptides: targeting protein-protein interactions in drug development. Explor Drug Sci. 2024;2(2):154–89. doi:10.37349/eds.2024.00041. [Google Scholar] [CrossRef]
118. Majewska M, Zamlynny V, Pieta IS, Nowakowski R, Pieta P. Interaction of LL-37 human cathelicidin peptide with a model microbial-like lipid membrane. Bioelectrochem. 2021;141(Pt 3):107842. doi:10.1016/j.bioelechem.2021.107842. [Google Scholar] [PubMed] [CrossRef]
119. Dabbagh Moghaddam F, Akbarzadeh I, Marzbankia E, Farid M, Khaledi L, Reihani AH, et al. Delivery of melittin-loaded niosomes for breast cancer treatment: an in vitro and in vivo evaluation of anti-cancer effect. Cancer Nanotechnol. 2021;12(1):14. doi:10.1186/s12645-021-00085-9. [Google Scholar] [CrossRef]
120. Javid H, Oryani MA, Rezagholinejad N, Esparham A, Tajaldini M, Karimi-Shahri M. RGD peptide in cancer targeting: benefits, challenges, solutions, and possible integrin-RGD interactions. Cancer Med. 2024;13(2):e6800. doi:10.1002/cam4.6800. [Google Scholar] [PubMed] [CrossRef]
121. Zuo H. iRGD: a promising peptide for cancer imaging and a potential therapeutic agent for various cancers. J Oncol. 2019;2019(1):9367845. doi:10.1155/2019/9367845. [Google Scholar] [PubMed] [CrossRef]
122. Lopes R, Shi K, Fonseca NA, Gama A, Ramalho JS, Almeida L, et al. Modelling the impact of nucleolin expression level on the activity of F3 peptide-targeted pH-sensitive pegylated liposomes containing doxorubicin. Drug Deliv Transl Res. 2022;2022(3):1–18. doi:10.1007/s13346-021-00972-z. [Google Scholar] [PubMed] [CrossRef]
123. Roscoe I, Parker M, Dong D, Li X, Li Z. Human serum albumin and the p53-derived peptide fusion protein promotes cytotoxicity irrespective of p53 status in cancer cells. Mol Pharm. 2018;15(11):5046–57. doi:10.1021/acs.molpharmaceut.8b00647. [Google Scholar] [PubMed] [CrossRef]
124. Luo G-F, Chen W-H, Liu Y, Lei Q, Zhuo R-X, Zhang X-Z. Multifunctional enveloped mesoporous silica nanoparticles for subcellular co-delivery of drug and therapeutic peptide. Sci Rep. 2014;4(1):6064. doi:10.1038/srep06064. [Google Scholar] [PubMed] [CrossRef]
125. Saraswathy SD, Mirunalini A, Karthikeyan K, Premkumar K. BH3 mimetic peptides: an effective strategy to complement anticancer therapy. Curr Protein Pept Sci. 2023;24(10):853–64. doi:10.2174/1389203724666230822100131. [Google Scholar] [PubMed] [CrossRef]
126. Jin W, Qin B, Chen Z, Liu H, Barve A, Cheng K. Discovery of PSMA-specific peptide ligands for targeted drug delivery. Int J Pharm. 2016;513(1–2):138–47. doi:10.1016/j.ijpharm.2016.08.048. [Google Scholar] [PubMed] [CrossRef]
127. Kim Y, Lillo AM, Steiniger SC, Liu Y, Ballatore C, Anichini A, et al. Targeting heat shock proteins on cancer cells: selection, characterization, and cell-penetrating properties of a peptidic GRP78 ligand. Biochem. 2006;45(31):9434–44. doi:10.1021/bi060264j. [Google Scholar] [PubMed] [CrossRef]
128. Hariharan S, Gustafson D, Holden S, McConkey D, Davis D, Morrow M, et al. Assessment of the biological and pharmacological effects of the ανβ3 and ανβ5 integrin receptor antagonist, cilengitide (EMD 121974in patients with advanced solid tumors. Ann Oncol. 2007;18(8):1400–7. doi:10.1093/annonc/mdm140. [Google Scholar] [PubMed] [CrossRef]
129. Essler M, Ruoslahti E. Molecular specialization of breast vasculature: a breast-homing phage-displayed peptide binds to aminopeptidase P in breast vasculature. Proc Natl Acad Sci. 2002;99(4):2252–7. doi:10.1073/pnas.251687998. [Google Scholar] [PubMed] [CrossRef]
130. Mueller J, Gaertner FC, Blechert B, Janssen K-P, Essler M. Targeting of tumor blood vessels: a phage-displayed tumor-homing peptide specifically binds to matrix metalloproteinase-2-processed collagen IV and blocks angiogenesis in vivo. Mol Cancer Res. 2009;7(7):1078–85. doi:10.1158/1541-7786.MCR-08-0538. [Google Scholar] [PubMed] [CrossRef]
131. Green DR. The mitochondrial pathway of apoptosis part II: the BCL-2 protein family. Cold Spring Harb Perspect Biol. 2022;14(6):a041046. doi:10.1101/cshperspect.a041046. [Google Scholar] [PubMed] [CrossRef]
132. Tung C, Varzideh F, Farroni E, Mone P, Kansakar U, Jankauskas SS, et al. Elamipretide: a review of its structure, mechanism of action, and therapeutic potential. Int J Mol Sci. 2025;26(3):944. doi:10.3390/ijms26030944. [Google Scholar] [PubMed] [CrossRef]
133. Peixoto PM, Lue JK, Ryu S-Y, Wroble BN, Sible JC, Kinnally KW. Mitochondrial apoptosis-induced channel (MAC) function triggers a Bax/Bak-dependent bystander effect. Am J Pathol. 2011;178(1):48–54. doi:10.1016/j.ajpath.2010.11.014. [Google Scholar] [PubMed] [CrossRef]
134. Wang L, Liu W-Q, Broussy S, Han B, Fang H. Recent advances of anti-angiogenic inhibitors targeting VEGF/VEGFR axis. Front Pharmacol. 2024;14:1307860. doi:10.3389/fphar.2023.1307860. [Google Scholar] [PubMed] [CrossRef]
135. El-Mousawi M, Tchistiakova L, Yurchenko L, Pietrzynski G, Moreno M, Stanimirovic D, et al. A vascular endothelial growth factor high affinity receptor 1-specific peptide with antiangiogenic activity identified using a phage display peptide library. J Biol Chem. 2003;278(47):46681–91. doi:10.1074/jbc.M308681200. [Google Scholar] [PubMed] [CrossRef]
136. Lu L, Chen H, Hao D, Zhang X, Wang F. The functions and applications of A7R in anti-angiogenic therapy, imaging and drug delivery systems. Asian J Pharm Sci. 2019;14(6):595–608. doi:10.1016/j.ajps.2019.04.004. [Google Scholar] [PubMed] [CrossRef]
137. Petreaca ML, Yao M, Liu Y, DeFea K, Martins-Green M. Transactivation of vascular endothelial growth factor receptor-2 by interleukin-8 (IL-8/CXCL8) is required for IL-8/CXCL8-induced endothelial permeability. Mol Biol Cell. 2007;18(12):5014–23. doi:10.1091/mbc.e07-01-0004. [Google Scholar] [PubMed] [CrossRef]
138. Kaur D, Deshmukh R. Physiology of cellular demise: apoptosis, necrosis, and autophagy. Clin Perspect Target Ther Apopt: Elsevier. 2021;14(11):23–78. doi:10.1016/B978-0-12-815762-6.00002-0. [Google Scholar] [CrossRef]
139. Srivastava RK. TRAIL/Apo-2L: mechanisms and clinical applications in cancer. Neoplasia. 2001;3(6):535–46. doi:10.1038/sj.neo.7900203. [Google Scholar] [PubMed] [CrossRef]
140. Schmidt SR. Fusion proteins: current status and future perspectives. In: Bioprocessing, bioengineering and process chemistry in the biopharmaceutical industry: using chemistry and bioengineering to improve the performance of biologics. Cham, Switzerland; 2024. p. 287–343. doi:10.1007/978-3-031-62007-2_11. [Google Scholar] [CrossRef]
141. Ruocco MG, Pilones KA, Kawashima N, Cammer M, Huang J, Babb JS, et al. Suppressing T cell motility induced by anti-CTLA-4 monotherapy improves antitumor effects. J Clin Invest. 2012;122(10):3718–30. doi:10.1172/JCI61931. [Google Scholar] [PubMed] [CrossRef]
142. Lischer CDA. Computational methods to find and rank MHC-I restricted tumor-associated antigens to improve therapeutic efficacy and tolerability of antigen-based cancer immunotherapy. Germany: Friedrich-Alexander-Universitaet Erlangen-Nuernberg; 2023. [Google Scholar]
143. Ren J, Li B. The functional stability of FOXP3 and RORγt in Treg and Th17 and their therapeutic applications. Adv Protein Chem Struct Biol. 2017;107(8):155–89. doi:10.1016/bs.apcsb.2016.10.002. [Google Scholar] [PubMed] [CrossRef]
144. Goyal P, Malviya R. Advances in nuclei targeted delivery of nanoparticles for the management of cancer. BBA Rev Cancer. 2023;1878(3):188881. doi:10.1016/j.bbcan.2023.188881. [Google Scholar] [PubMed] [CrossRef]
145. Frolova AS, Chepikova OE, Deviataikina AS, Solonkina AD, Zamyatnin AAJr. New perspectives on the role of nuclear proteases in cell death pathways. Biol. 2023;12(6):797. doi:10.3390/biology12060797. [Google Scholar] [PubMed] [CrossRef]
146. Herman JA. Investigation of the molecular mechanisms and therapeutic potential of oncogene-induced kinetochore-microtubule defects. Fort Collins, CO, USA: Colorado State University; 2015. [Google Scholar]
147. Eike L-M, Yang N, Rekdal Ø, Sveinbjørnsson B. The oncolytic peptide LTX-315 induces cell death and DAMP release by mitochondria distortion in human melanoma cells. Oncotarget. 2015;6(33):34910–23. doi:10.18632/oncotarget.5308. [Google Scholar] [PubMed] [CrossRef]
148. Patel KD, De Zoysa GH, Kanamala M, Patel K, Pilkington LI, Barker D, et al. Novel cell-penetrating peptide conjugated proteasome inhibitors: anticancer and antifungal investigations. J Med Chem. 2019;63(1):334–48. doi:10.1021/acs.jmedchem.9b01694. [Google Scholar] [PubMed] [CrossRef]
149. Zhong S, Jeong J-H, Chen Z, Chen Z, Luo J-L. Targeting tumor microenvironment by small-molecule inhibitors. Transl Oncol. 2020;13(1):57–69. doi:10.1016/j.tranon.2019.10.001. [Google Scholar] [PubMed] [CrossRef]
150. Dharmaratne NU, Kaplan AR, Glazer PM. Targeting the hypoxic and acidic tumor microenvironment with pH-sensitive peptides. Cells. 2021;10(3):541. doi:10.3390/cells10030541. [Google Scholar] [PubMed] [CrossRef]
151. Huang J, Zhang L, Wan D, Zhou L, Zheng S, Lin S, et al. Extracellular matrix and its therapeutic potential for cancer treatment. Signal Transduct Target Ther. 2021;6(1):153. doi:10.1038/s41392-021-00544-0. [Google Scholar] [PubMed] [CrossRef]
152. Mitchell MJ, Billingsley MM, Haley RM, Wechsler ME, Peppas NA, Langer R. Engineering precision nanoparticles for drug delivery. Nat Rev Drug Discov. 2021;20(2):101–24. doi:10.1038/s41573-020-0090-8. [Google Scholar] [PubMed] [CrossRef]
153. Nikitovic D, Kukovyakina E, Berdiaki A, Tzanakakis A, Luss A, Vlaskina E, et al. Enhancing tumor targeted therapy: the role of iRGD peptide in advanced drug delivery systems. Cancers. 2024;16(22):3768. doi:10.3390/cancers16223768. [Google Scholar] [PubMed] [CrossRef]
154. Souri M, Soltani M, Kashkooli FM, Shahvandi MK. Engineered strategies to enhance tumor penetration of drug-loaded nanoparticles. J Control Release. 2022;341:227–46. doi:10.1016/j.jconrel.2021.11.024. [Google Scholar] [PubMed] [CrossRef]
155. Wang A, Zheng Y, Zhu W, Yang L, Yang Y, Peng J. Melittin-based nano-delivery systems for cancer therapy. Biomolecules. 2022;12(1):118. doi:10.3390/biom12010118. [Google Scholar] [PubMed] [CrossRef]
156. Sonju JJ, Dahal A, Singh SS, Jois SD. Peptide-functionalized liposomes as therapeutic and diagnostic tools for cancer treatment. J Control Release. 2021;329:624–44. doi:10.1016/j.jconrel.2020.09.055. [Google Scholar] [PubMed] [CrossRef]
157. Liu P, Chen G, Zhang J. A review of liposomes as a drug delivery system: current status of approved products, regulatory environments, and future perspectives. Molecules. 2022;27(4):1372. doi:10.3390/molecules27041372. [Google Scholar] [PubMed] [CrossRef]
158. Saini L, Dubey A, Pal R, Pandey P, Mandal RK. Synthetic and natural polymers enhancing drug delivery and their treatment: a comprehensive review. J Drug Deliv Ther. 2024;14(10):153–65. doi:10.22270/jddt.v14i10.6802. [Google Scholar] [CrossRef]
159. Ding L, Agrawal P, Singh SK, Chhonker YS, Sun J, Murry DJ. Polymer-based drug delivery systems for cancer therapeutics. Polym. 2024;16(6):843. doi:10.3390/polym16060843. [Google Scholar] [PubMed] [CrossRef]
160. Khaliq NU, Lee J, Kim S, Sung D, Kim H. Pluronic F-68 and F-127 based nanomedicines for advancing combination cancer therapy. Pharmaceutics. 2023;15(8):2102. doi:10.3390/pharmaceutics15082102. [Google Scholar] [PubMed] [CrossRef]
161. Hughes DL. Patent review of manufacturing routes to oncology drugs: carfilzomib, osimertinib, and venetoclax. Org Process Res Dev. 2016;20(12):2028–42. doi:10.1021/acs.oprd.6b00374. [Google Scholar] [CrossRef]
162. Manoharan P, Lamarca A, Navalkissoor S, Calero J, San Chan P, Julyan P, et al. Safety, tolerability and clinical implementation of ‘ready-to-use’ 68gallium-DOTA0-Tyr3-octreotide (68Ga-DOTATOC)(SomaKIT TOC) for injection in patients diagnosed with gastroenteropancreatic neuroendocrine tumours (GEP-NETs). ESMO Open. 2020;5(2):e000650. doi:10.1136/esmoopen-2019-000650. [Google Scholar] [PubMed] [CrossRef]
163. Deeks ED. Venetoclax: first global approval. Drugs. 2016;76(9):979–87. doi:10.1007/s40265-016-0596-x. [Google Scholar] [PubMed] [CrossRef]
164. Greig SL. Osimertinib: first global approval. Drugs. 2016;76(2):263–73. doi:10.1007/s40265-015-0533-4. [Google Scholar] [PubMed] [CrossRef]
165. Ma X, Tian Y, Yang R, Wang H, Allahou LW, Chang J, et al. Nanotechnology in healthcare, and its safety and environmental risks. J Nanobiotechnol. 2024;22(1):715. doi:10.1186/s12951-024-02901-x. [Google Scholar] [PubMed] [CrossRef]
166. Song JS, Tawa M, Chau NG, Kupper TS, LeBoeuf NR. Aprepitant for refractory cutaneous T-cell lymphoma-associated pruritus: 4 cases and a review of the literature. BMC Cancer. 2017;17(1):1–6. doi:10.1186/s12885-017-3194-8. [Google Scholar] [PubMed] [CrossRef]
167. Ranasinghe R, Mathai ML, Zulli A. Cisplatin for cancer therapy and overcoming chemoresistance. Heliyon. 2022;8(9):e10608. doi:10.1016/j.heliyon.2022.e10608. [Google Scholar] [PubMed] [CrossRef]
168. Bardia A, Hurvitz SA, Tolaney SM, Loirat D, Punie K, Oliveira M, et al. Sacituzumab govitecan in metastatic triple-negative breast cancer. New England J Med. 2021;384(16):1529–41. doi:10.1056/NEJMoa2028485. [Google Scholar] [PubMed] [CrossRef]
169. Sartor O, De Bono J, Chi KN, Fizazi K, Herrmann K, Rahbar K, et al. Lutetium-177-PSMA-617 for metastatic castration-resistant prostate cancer. New England J Med. 2021;385(12):1091–103. doi:10.1056/NEJMoa2107322. [Google Scholar] [PubMed] [CrossRef]
170. Olsen E, Duvic M, Frankel A, Kim Y, Martin A, Vonderheid E, et al. Pivotal phase III trial of two dose levels of denileukin diftitox for the treatment of cutaneous T-cell lymphoma. J Clin Oncol. 2001;19(2):376–88. doi:10.1200/JCO.2001.19.2.376. [Google Scholar] [PubMed] [CrossRef]
171. Rudin CM, Pietanza MC, Bauer TM, Ready N, Morgensztern D, Glisson BS, et al. Rovalpituzumab tesirine, a DLL3-targeted antibody-drug conjugate, in recurrent small-cell lung cancer: a first-in-human, first-in-class, open-label, phase 1 study. Lancet Oncol. 2017;18(1):42–51. doi:10.1016/S1470-2045(16)30565-4. [Google Scholar] [PubMed] [CrossRef]
172. Tabernero J, Yoshino T, Cohn AL, Obermannova R, Bodoky G, Garcia-Carbonero R, et al. Ramucirumab versus placebo in combination with second-line FOLFIRI in patients with metastatic colorectal carcinoma that progressed during or after first-line therapy with bevacizumab, oxaliplatin, and a fluoropyrimidine (RAISEa randomised, double-blind, multicentre, phase 3 study. Lancet Oncol. 2015;16(5):499–508. doi:10.1016/S1470-2045(15)70127-0. [Google Scholar] [PubMed] [CrossRef]
173. Mirdita M, Schütze K, Moriwaki Y, Heo L, Ovchinnikov S, Steinegger M. ColabFold: making protein folding accessible to all. Nat Met. 2022;19(6):679–82. doi:10.1038/s41592-022-01488-1. [Google Scholar] [PubMed] [CrossRef]
174. Waterhouse A, Bertoni M, Bienert S, Studer G, Tauriello G, Gumienny R, et al. SWISS-MODEL: homology modelling of protein structures and complexes. Nucleic Acids Res. 2018;46(W1):W296–303. doi:10.1093/nar/gky427. [Google Scholar] [PubMed] [CrossRef]
175. Alqurashi A. Bioinformatic analysis of oxygen sensitivity in Alorcobacter bivalviorum’s pyruvate: ferredoxin oxidoreductase (PFOR). Adv Life Sci. 2024;11(4):843–8. doi:10.62940/als.v11i4.3142. [Google Scholar] [CrossRef]
176. Imani S, Li X, Chen K, Maghsoudloo M, Kaboli PJ, Hashemi M, et al. Computational biology and artificial intelligence in mRNA vaccine design for cancer immunotherapy. Front Cell Infect Microbiol. 2025;14:1501010. doi:10.3389/fcimb.2024.1501010. [Google Scholar] [PubMed] [CrossRef]
177. Yang S, Kar S, Leszczynski J. Tools and software for computer-aided drug design and discovery. In: Cheminformatics, QSAR and machine learning applications for novel drug development. Elsevier; 2023. p. 637–61. doi:10.1016/B978-0-443-18638-7.00017-7. [Google Scholar] [CrossRef]
178. Du Z, Su H, Wang W, Ye L, Wei H, Peng Z, et al. The trRosetta server for fast and accurate protein structure prediction. Nat Prot. 2021;16(12):5634–51. doi:10.1038/s41596-021-00628-9. [Google Scholar] [PubMed] [CrossRef]
179. Laskowski R, MacArthur M, Thornton J. PROCHECK: validation of protein-structure coordinates; 2006 [cited 2025 Feb 10]. Available from: https://onlinelibrary.wiley.com/iucr/itc/Fa/ch25o2v0001/sec25o2o6/. [Google Scholar]
180. Alshehri AA, Almutairi AM, Shafie A, Alshehri NA, Almutairi SM, Anjum F. Identification of potential inhibitors targeting DNA adenine methyltransferase of Klebsiella pneumoniae for antimicrobial resistance management: a structure-based molecular docking study. Adv Life Sci. 2024;10(4):604–8. [Google Scholar]
181. Chen VB, Arendall WB, Headd JJ, Keedy DA, Immormino RM, Kapral GJ, et al. MolProbity: all-atom structure validation for macromolecular crystallography. Acta Crystallographica Section D: Biol Crystal. 2010;66(1):12–21. doi:10.1107/S0907444909042073. [Google Scholar] [PubMed] [CrossRef]
182. Benkert P, Künzli M, Schwede T. QMEAN server for protein model quality estimation. Nucleic Acids Res. 2009;37(suppl_2):W510–4. doi:10.1093/nar/gkp322. [Google Scholar] [PubMed] [CrossRef]
183. Javaid MS, Kaul H, Fazal N, Yaqub F, Naseer N, Hanif M, et al. In silico analysis to reveal underlying trans differentiation mechanism of mesenchymal stem cells into osteocytes. Adv Life Sci. 2021;8(4):412–8. [Google Scholar]
184. Eisenberg D, Lüthy R, Bowie JU. [20] VERIFY3D: assessment of protein models with three-dimensional profiles. In: Methods in Enzymology. Amsterdam, Netherlands: Elsevier; 1997. p. 396–404. doi:10.1016/S0076-6879(97)77022-8. [Google Scholar] [CrossRef]
185. Ghani U, Desta I, Jindal A, Khan O, Jones G, Hashemi N, et al. Improved docking of protein models by a combination of Alphafold2 and ClusPro. BioRxiv. 2021. doi:10.1101/2021.09.07.459290. [Google Scholar] [CrossRef]
186. Honorato RV, Trellet ME, Jiménez-García B, Schaarschmidt JJ, Giulini M, Reys V, et al. The HADDOCK2. 4 web server for integrative modeling of biomolecular complexes. Nat Protoc. 2024;19(11):3219–41. doi:10.1038/s41596-024-01011-0. [Google Scholar] [PubMed] [CrossRef]
187. Muhammad Rehman H, Rehman HM, Naveed M, Khan MT, Shabbir MA, Aslam S, et al. In silico investigation of a chimeric IL24-LK6 fusion protein as a potent candidate against breast cancer. Bioinform Biol Insights. 2023;17:1–13. doi:10.1177/11779322231182560. [Google Scholar] [PubMed] [CrossRef]
188. Pierce BG, Wiehe K, Hwang H, Kim B-H, Vreven T, Weng Z. ZDOCK server: interactive docking prediction of protein-protein complexes and symmetric multimers. Bioinform. 2014;30(12):1771–3. doi:10.1093/bioinformatics/btu097. [Google Scholar] [PubMed] [CrossRef]
189. Imran M, Alshammari AK, Alfarh KF, Alanazi RK, Alfrah SF, Alshammri OM, et al. In-vitro and in-silico studies based discovery of 2-aryl-N-(4-morpholinophenyl) thiazol-4-amines as promising DNA gyrase inhibitors. Adv Life Sci. 2024;11(4):912–7. doi:10.62940/als.v11i4.3514. [Google Scholar] [CrossRef]
190. Mursal M, Ahmad M, Hussain S, Khan MF. Navigating the computational seas: a comprehensive overview of molecular docking software in drug discovery. 2024. doi:10.5772/intechopen.1004802. [Google Scholar] [CrossRef]
191. Pettersen EF, Goddard TD, Huang CC, Couch GS, Greenblatt DM, Meng EC, et al. UCSF Chimera—a visualization system for exploratory research and analysis. J Comput Chem. 2004;25(13):1605–12. doi:10.1002/jcc.20084. [Google Scholar] [PubMed] [CrossRef]
192. Saif R, Ashfaq K, Ali G, Iftekhar A, Zia S, Yousaf MZ. Computational prediction of Cassia angustifolia compounds as a potential drug agents against main protease of SARS-nCov2. Adv Life Sci. 2022;9(1):36–40. [Google Scholar]
193. Andersson MI, Murugan NA, Podobas A, Markidis S, editors. Breaking down the parallel performance of gromacs, a high-performance molecular dynamics software. In: International Conference on Parallel Processing and Applied Mathematics; 2022; Springer. doi:10.1007/978-3-031-30442-2_25. [Google Scholar] [CrossRef]
194. Barhaghi MS, Crawford B, Schwing G, Hardy DJ, Stone JE, Schwiebert L, et al. Py-MCMD: python software for performing hybrid Monte Carlo/molecular dynamics simulations with GOMC and NAMD. J Chem The Comput. 2022;18(8):4983–94. doi:10.1021/acs.jctc.1c00911. [Google Scholar] [PubMed] [CrossRef]
195. Wang J, Wolf RM, Caldwell JW, Kollman PA, Case DA. Development and testing of a general amber force field. J Computat Chem. 2004;25(9):1157–74. doi:10.1002/jcc.20035. [Google Scholar] [PubMed] [CrossRef]
196. Asiri A. In-silico perspectives on the potential therapeutic aids of hesperetin derivatives for lung cancer. Adv Life Sci. 2024;11(4):878–86. doi:10.62940/als.v11i4.3308. [Google Scholar] [CrossRef]
197. Rehman HM, Yousaf N, Hina SM, Nadeem T, Ansari MA, Chaudry A, et al. Design and computational analysis of a novel Azurin-BR2 chimeric protein against breast cancer. Toxicol Res. 2024;13(6):tfae179. doi:10.1093/toxres/tfae179. [Google Scholar] [PubMed] [CrossRef]
198. Khalid S, Rehman HM, Al-Qassab Y, Ahmad I, Fatima T, Mubasher MM, et al. Design and computational analysis of a novel Leptulipin-p28 fusion protein as a multitarget anticancer therapy in breast cancer. Toxicol Res. 2024;13(5):tfae174. doi:10.1093/toxres/tfae174. [Google Scholar] [PubMed] [CrossRef]
199. Rehman HM, Rehman HM, Ahmed N, Imran M. In silico Design and Evaluation of Novel Cell Targeting Melittin-Interleukin-24 Fusion Protein: a Potential Drug Candidate Against Breast Cancer. Sains Malaysiana. 2023;52(11):3223–37. doi:10.17576/jsm-2023-5211-15. [Google Scholar] [CrossRef]
200. Aslam S, Rehman HM, Sarwar MZ, Ahmad A, Ahmed N, Amirzada MI, et al. Computational modeling, high-level soluble expression and in vitro cytotoxicity assessment of recombinant Pseudomonas aeruginosa azurin: a promising anti-cancer therapeutic candidate. Pharmaceutics. 2023;15(7):1825. doi:10.3390/pharmaceutics15071825. [Google Scholar] [PubMed] [CrossRef]
201. Qureshi S, Ahmed N, Rehman HM, Amirzada MI, Saleem F, Waheed K, et al. Investigation of therapeutic potential of the Il24-p20 fusion protein against breast cancer: an in-silico approach. Silico Pharmacol. 2024;12(2):84. doi:10.1007/s40203-024-00252-x. [Google Scholar] [PubMed] [CrossRef]
202. Fatima T, Mubasher MM, Rehman HM, Niyazi S, Alanzi AR, Kalsoom M, et al. Computational modeling study of IL-15-NGR peptide fusion protein: a targeted therapeutics for hepatocellular carcinoma. AMB Exp. 2024;14(1):91. doi:10.1186/s13568-024-01747-8. [Google Scholar] [PubMed] [CrossRef]
203. Hafiz Muhammad R, Wardah S, Muhammad Naveed K, Numan Y, Fareeha B, Hamid B, et al. A comprehensive in silico study of the NDB-IL-24 fusion protein for tumor targeting: a promising anti-cancer therapeutic candidate. J Bio Reg Home Agents. 2024;38(4):3449–61. doi:10.23812/j.biol.regul.homeost.agents.20243804.274. [Google Scholar] [CrossRef]
204. Moghadam ZM, Halabian R, Sedighian H, Behzadi E, Amani J, Fooladi AAI. Designing and analyzing the structure of DT-STXB fusion protein as an anti-tumor agent: an in silico approach. Iranian J Pathol. 2019;14(4):305–12. doi:10.30699/IJP.2019.101200.2004. [Google Scholar] [PubMed] [CrossRef]
205. Goleij Z, Hosseini HM, Amin M, Amani J, Behzadi E, Fooladi AAI. In silico evaluation of two targeted chimeric proteins based on bacterial toxins for breast cancer therapy. Int J Cancer Manag. 2019;12(2):e83315. doi:10.5812/ijcm.83315. [Google Scholar] [CrossRef]
Cite This Article
 Copyright © 2025 The Author(s). Published by Tech Science Press.
Copyright © 2025 The Author(s). Published by Tech Science Press.This work is licensed under a Creative Commons Attribution 4.0 International License , which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
 Downloads
Downloads
 Citation Tools
Citation Tools