
 Submit a Paper
Submit a Paper Propose a Special lssue
Propose a Special lssue Open Access
Open Access
ARTICLE

Combining Ability and Heterotic Effects in Newly Developed Early Maturing and High-Yielding Maize Hybrids under Low and Recommended Nitrogen Conditions
1 Department of Agronomy, Faculty of Agriculture, Kafrelsheikh University, Kafr El-Sheikh, 33516, Egypt
2 Department of Biology, Faculty of Science, Umm Al-Qura University, Makkah, 24372, Saudi Arabia
3 Department of Biological Sciences, Faculty of Science, King Abdulaziz University, Jeddah, 21589, Saudi Arabia
4 Department of Biological Science, College of Science, University of Jeddah, Jeddah, 21589, Saudi Arabia
5 Department of Biology, College of Science, Buraydah, 51452, Saudi Arabia
6 Department of Plant Production, Genetic Branch, Faculty of Environmental and Agricultural Sciences, Arish University, El-Arish, 45511, Egypt
7 Department of Geography and Environmental Sustainability, College of Humanities and Social Sciences, Princess Nourah bint Abdulrahman University, P.O. Box 84428, Riyadh, 11671, Saudi Arabia
* Corresponding Author: Mohamed M. Kamara. Email:
(This article belongs to the Special Issue: Integrative Approaches to Plant Stress Responses under Changing Climate Conditions)
Phyton-International Journal of Experimental Botany 2025, 94(1), 101-122. https://doi.org/10.32604/phyton.2025.058033
Received 09 September 2024; Accepted 07 November 2024; Issue published 24 January 2025
 View Full Text
View Full Text Download PDF
Download PDFAbstract
Nitrogen (N) is a crucial nutrient vital for the growth and productivity of maize. However, excessive nitrogen application can result in numerous environmental and ecological problems, such as water pollution, biodiversity loss, and greenhouse gas emissions. Therefore, breeding maize hybrids resilient to low nitrogen conditions is crucial for sustainable agriculture, especially under low nitrogen conditions. Consequently, this study aimed to evaluate the combining ability and heterosis of maize lines, recognize promising hybrids, and study gene action controlling key traits under low and recommended N stress conditions. The half-diallel mating design hybridized seven maize inbreds, resulting in 21 F1 hybrids. These hybrids, along with two high-yielding commercial hybrids (SC10 and TWC310), were evaluated in field trials under recommended (290 kg/ha) and low N (166 kg N/ha) conditions. Significant variations were observed among assessed hybrids for all measured traits, with non-additive gene action being predominant for grain yield and its related characteristics under recommended and low N conditions. Inbred lines P105 and P106 were recognized as effective combiners for earliness, with P105 also excelling in shorter plant height and lower ear placement. In addition, P101, P102, and P104 were identified as good combiners for increasing grain yield and related attributes under low N conditions. The crosses P105 × P106 and P106 × P107 demonstrated outstanding heterotic effects for earliness, while hybrids P101 × P102 and P102 × P104 exhibited remarkable heterotic effects for grain yield low nitrogen stress conditions. These promising hybrids could be considered for commercial use after further evaluation. Strong positive correlations were found between grain yield and ear height, plant height, number of kernels per row, and 1000-grain weight, highlighting their importance for indirect selection to enhance the grain yield of maize under low N stress conditions.Keywords
Maize (Zea mays) plays a vital role globally, being a key source of nutrition, livestock feed, and biofuel [1]. It is the third most produced crop worldwide, following wheat and rice. It is cultivated on about 203.5 million hectares, yielding 1163.5 million tons of grain [2]. Maize is a significant source of national income in numerous countries [3]. Despite its global importance, Egypt faces a notable gap between maize production and consumption. This gap is primarily driven by the country’s limited arable land, water scarcity, and growing population, which collectively constrain domestic maize production. Therefore, maize breeders are focusing on developing new high-yielding maize hybrids to harness heterosis and identify the most promising varieties for commercial production.
Conventional nitrogen fertilizers can result in environmental problems like water contamination and the release of greenhouse gases [4]. Therefore, breeding maize hybrids under limited nitrogen is crucial to reducing dependency on these inputs. Such hybrids enhance soil health by minimizing excessive fertilizer use and contributing to higher agricultural productivity in challenging soil conditions [5]. Improving maize’s ability to grow and yield effectively in low nitrogen environments can promote more sustainable farming practices, ensure food security, and support the long-term health of agricultural ecosystems.
Early flowering in maize offers significant benefits, including water conservation and enhanced resistance to specific pests. By reducing the time to flowering, maize plants can minimize their exposure to pest infestations, such as those caused by the African pink borer (Sesamia cretica), the pink spotted borer (Pyrausta nubilialis), and the spotted borer (Chilo simplex). This shortened developmental period helps to mitigate the impact of these damaging pests, which are more likely to infest crops during prolonged growth stages. Consequently, early flowering can lead to more efficient use of irrigation resources and contribute to better overall crop protection and yield stability. Heterosis, or hybrid vigor, is a pivotal concept in plant breeding, offering the potential to develop maize hybrids with higher yields and early maturing hybrids under normal and stressed environments [6]. The application of heterosis is crucial for classifying maize lines into specific heterotic groups, which aids in creating high-performing hybrids with superior agronomic traits compared to existing commercial varieties. Additionally, exploiting hybrid vigor is vital for advancing sustainable agriculture by reducing the impacts of environmental stress and improving food production efficiency to meet growing global demands.
Studying the combining ability of maize inbred lines is pivotal for advancing maize breeding programs and improving crop performance. By assessing how different inbred lines interact when crossed, researchers can identify which parental combinations produce hybrids with enhanced traits such as higher yield, better disease resistance, or improved adaptability to varying environmental conditions [7]. This knowledge allows breeders to strategically select and combine inbred lines to harness the full potential of hybrid vigor or heterosis, resulting in more productive and resilient maize varieties [8,9]. Combining ability accelerates the development of high-performing hybrids and contributes to achieving greater agricultural sustainability and food security by optimizing crop performance under diverse growing conditions [10,11]. Moreover, understanding the genetic mechanisms underlying agronomic traits, such as yield characters, can pinpoint the specific genes and their interactions contributing to desirable characteristics. This knowledge enables breeders to make well-informed decisions by identifying parental lines and planning breeding programs, thereby enhancing the efficiency of developing new maize hybrids with targeted traits [7]. Accordingly, this study aimed to (1) assess general combining ability of the assessed inbred lines and the specific combining ability of their hybrids under recommended and low nitrogen (N) conditions; (2) elucidate the inheritance patterns of the studied traits; (3) identify maize hybrids that exhibit high yield and early maturity under recommended and low N conditions; and (4) evaluate the relationships among the assessed traits under low N conditions.
2.1 Parental Lines and Hybridization
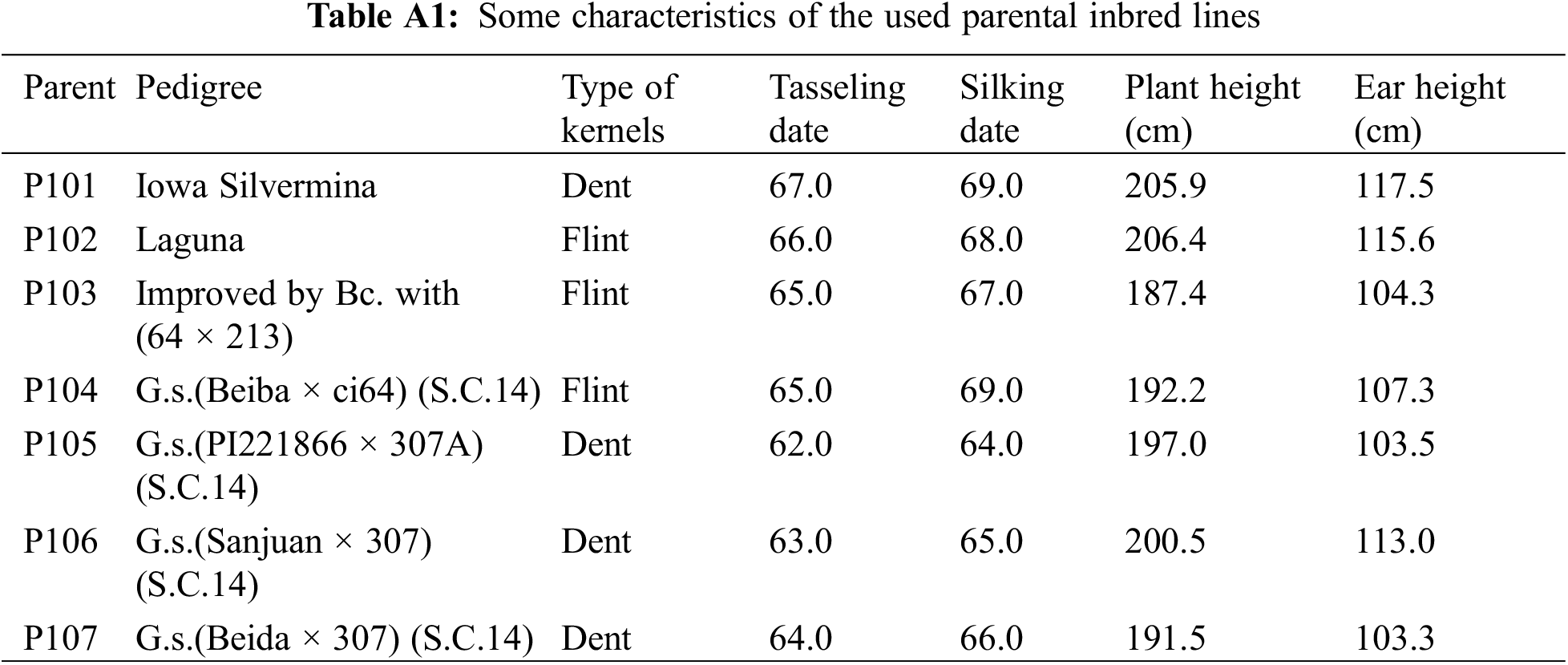
In this study, seven diverse white maize (Zea mays L.) inbred lines were utilized (Table A1), namely P101 (Inb. 7), P102 (Inb. 11), P103 (Inb. 37), P104 (Inb. 56), P105 (Inb. 81), P106 (Inb. 84), and P107 (Inb. 96). These inbred lines were obtained from Agricultural Research Center, Egypt. During the summer of 2021, the seeds of these inbred lines were divided and established on three different sowing dates (May 14, 21, and 28) to ensure sufficient hybrid seed production. The parental lines were sown in rows measuring 6 m in length with a spacing of 0.70 m between rows. Within each row, two seeds were sown per hill with a spacing of 0.25 m. The seedlings were thinned to one per hill after full emergence and before the first irrigation. A half-diallel mating design, excluding reciprocal crosses, was employed to create all possible cross combinations among the seven inbred lines, developing 21 F1 hybrids.
2.2 Field Trials and Recorded Traits
In the 2022 growing season, the generated F1 hybrids were evaluated alongside two high-yielding commercial check hybrids Single-Cross 10 (SC-10) and Three-Way Cross 310 (TWC-310) under recommended N (290 kg/ha) and low N stress conditions (166 kg N/ha) in a private farm, Kafr El-Sheikh, Egypt (31°56′N, 30°65′E). Meteorological data for both seasons of 2021 and 2022 are presented in Fig. 1. Soil samples were collected from 0–30 cm depths and 30–60 cm before sowing to analyze some chemical and physical properties. The analysis indicated that the soil was consistently clay throughout the entire profile, consisting of 51.79% clay, 23.84% silt, and 24.37% sand, with a bulk density of 1.38 g cm−3. The soil contained 1.55% organic matter, pH of 8.0, an electrical conductivity of 2.88 dS/m, and 4.4% calcium carbonate. The available nutrients were 30.1 mg N, 10.1 mg P, and 650 mg K per kg of soil. The soluble cations and anions were 112.3, 13.0, 30.1, 0.6, 22.7, 4.0, and 29.3 meq L−1 for Mg2+, Ca2+, N+, K+, Cl‒, HCO3−, and SO42−, respectively. A randomized Complete Block Design in three replicates was implemented. Each plot comprised two rows measuring 6 m in length and 0.7 m in width. Nitrogen fertilizer was added in two equal doses: the first dose was applied before the first irrigation, and the second dose was added before the second irrigation. Phosphorus fertilizer was incorporated during seedbed preparation at a rate of 70 kg/ha, using calcium superphosphate containing 15.5% P2O5. Additionally, 110 kg/ha of potassium sulfate with 48% K2O was applied after thinning.
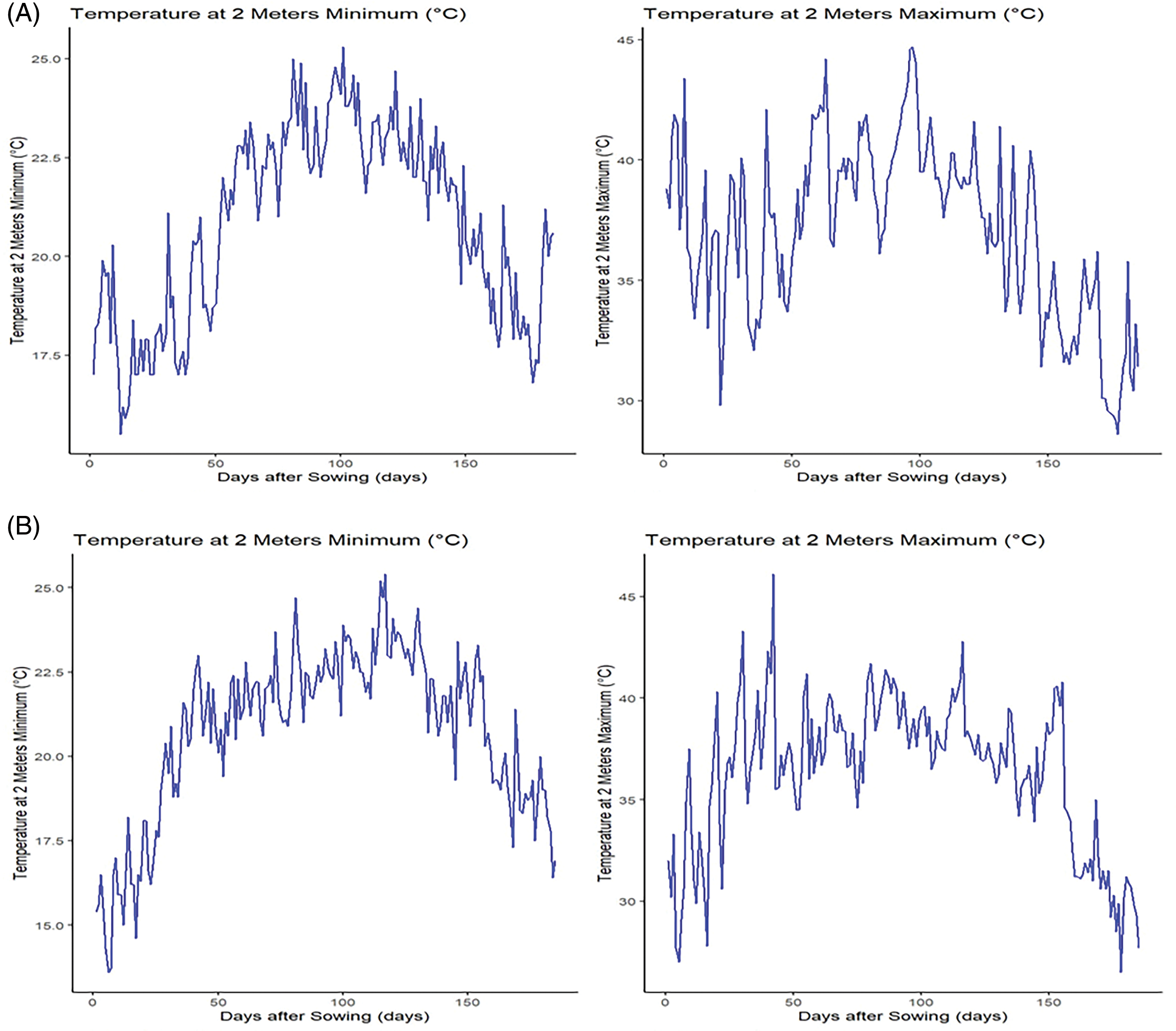
Figure 1: Meteorological data for the first season of 2021 (A) and the second season of 2022 (B)
Days to tasseling were recorded when 50% of the plants in each plot began producing tassels, and days to silking were noted when 50% of the plants in each plot started producing silks. Plant height was measured in centimeters from the ground to the top of the first tassel branch, and ear height was recorded as the distance from the ground to the base of the highest ear. During harvest, ten ears were randomly selected from each plot to assess agronomic traits, such as ear diameter, the number of kernels per row, and the weight of 1000 kernels. The plots were hand-harvested, and the weight of the shelled grain, adjusted to 15.5% moisture content, was utilized to estimate grain yield in tons per hectare.
Analysis of variance (ANOVA) was conducted for all traits, and the least significant difference (LSD) test (p < 0.05) was used to clarify the significance of differences among the assessed genotypes. Griffing’s method 4, model 1, was applied for combining ability analysis [12]. The statistical significance of the differences observed between the general combining ability (GCA) and specific combining ability (SCA) effects was determined by considering the standard error and the tabulated t-value. Standard heterosis was calculated by comparing the mean of the F1 hybrids to the two standard check (SC) hybrids using the formula: Heterosis over the standard check = [(F1 − SC)/SC] × 100. Principal component and heatmap analyses were conducted using R programming version 4.2.1.
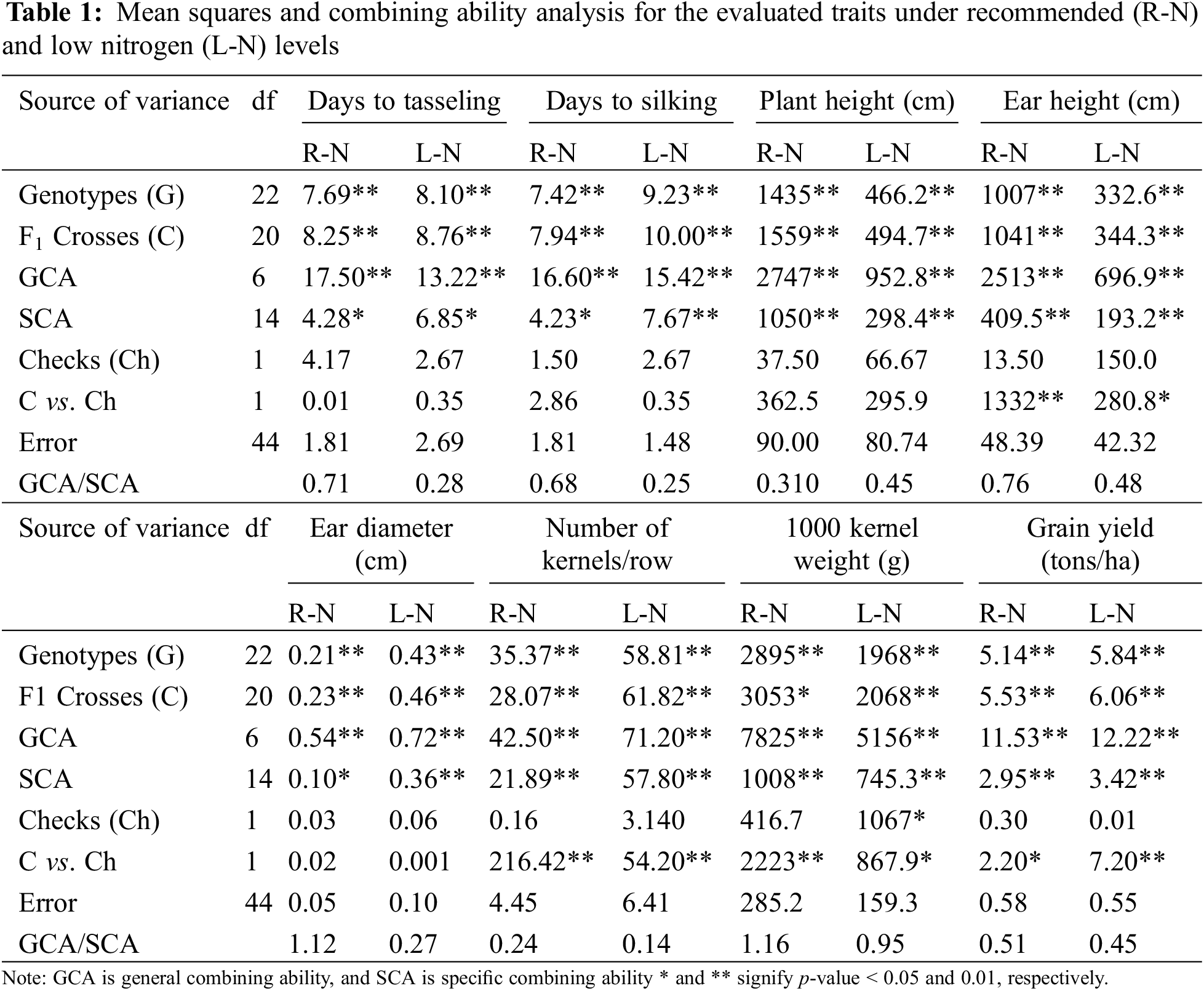
The mean squares for assessed genotypes were divided into components for crosses, checks, and the comparison between crosses and checks. Mean squares of both crosses and genotypes were highly significant for all evaluated traits under recommended and low N conditions (Table 1). However, the differences between the assessed check hybrids were insignificant across the studied characteristics under both conditions. Significant mean squares were detected for crosses vs. checks in traits including ear height, number of kernels per row, thousand kernel weight, and grain yield under both conditions. Additionally, variances due to GCA and SCA were highly significant for all traits under both recommended and low N conditions. The GCA/SCA ratio was less than one for all traits assessed under both conditions, except for ear diameter and thousand-kernel weight under normal conditions.
3.2 Performance of Assessed Hybrids
Nitrogen levels substantially impacted all measured traits, and low nitrogen stress significantly reduced all the measured traits. In comparison to the recommended nitrogen level, low N conditions significantly decreased days to tasseling by 6.12%, days to silking by 6.7%, plant height by 24.1%, ear height by 26.3%, ear diameter by 11.2%, number of kernels per row by 18.3%, thousand-kernel weight by 19.5%, and grain yield by 34.8% (Fig. 2). Significant variation was observed among the tested hybrids under recommended and low N conditions for each trait. The days to tasseling ranged from 58.7 days (P103 × P105) to 64.0 days (P103 × P104), with an average of 61.5 days under recommended conditions, while under low N it ranged from 54.0 days (P106 × P107) to 61.0 days (P103 × P104) with means of 57.8 days (Fig. 3A). Days to silking averaged 65.6 days under recommended conditions, fluctuating from 63.0 (P105 × P106) to 69.0 (P103 × P104), and 61.1 days under low N, ranging from 56.7 (P105 × P106) to 64.3 (P103 × P106) (Fig. 3B). For plant height, the range was from 203.3 cm (P106 × P107) to 288.3 cm (P101 × P107), with an average height of 262.7 cm under recommended conditions (Fig. 3C). In contrast, under low N conditions, the range was from 178.3 cm (P101 × P103) to 233.7 cm (P102 × P104), with an average height of 199.3 cm. Ear height averaged 150.4 cm, ranging from 111.3 cm (P106 × P107) to 174.7 cm (P102 × P103) under recommended conditions (Fig. 3D). In low N conditions, the average was 110.5 cm, ranging from 90.0 cm (P105 × P107) to 130.0 cm (P101 × P102). Ear diameter had an average of 4.89 cm under recommended and 4.34 cm under low N conditions, ranging from 4.43 (P105 × P107) to 5.47 cm (P104 × P106) and 3.30 cm (P105 × P107) to 4.90 cm (P102 × P104), respectively (Fig. 4A). Number of rows per ear, under recommended conditions, ranged from 36.6 (P105 × P107) to 43.6 (P101 × P102), and from 28.4 (P102 × P107) to 44.4 (P101 × P102) under stress conditions (Fig. 4B). Thousand-kernel weight differed from 280.0 to 410.0 g (average 354.9 g) under recommended conditions and from 222.0 to 335.0g (average 286.1 g) under low N, with (P103 × P104) showing the heaviest, and (P105 × P107) the lightest weights under both conditions (Fig. 4C). Grain yield varied between 5.89 tons/ha (P105 × P107) and 11.2 tons/ha (P101 × P102), with a mean yield of 8.40 tons/ha under optimal conditions (Fig. 4D). In contrast, under low nitrogen conditions, the yield ranged from 2.62 tons/ha (P105 × P107) to 8.17 tons/ha (P102 × P104), with an average yield of 5.42 tons/ha.
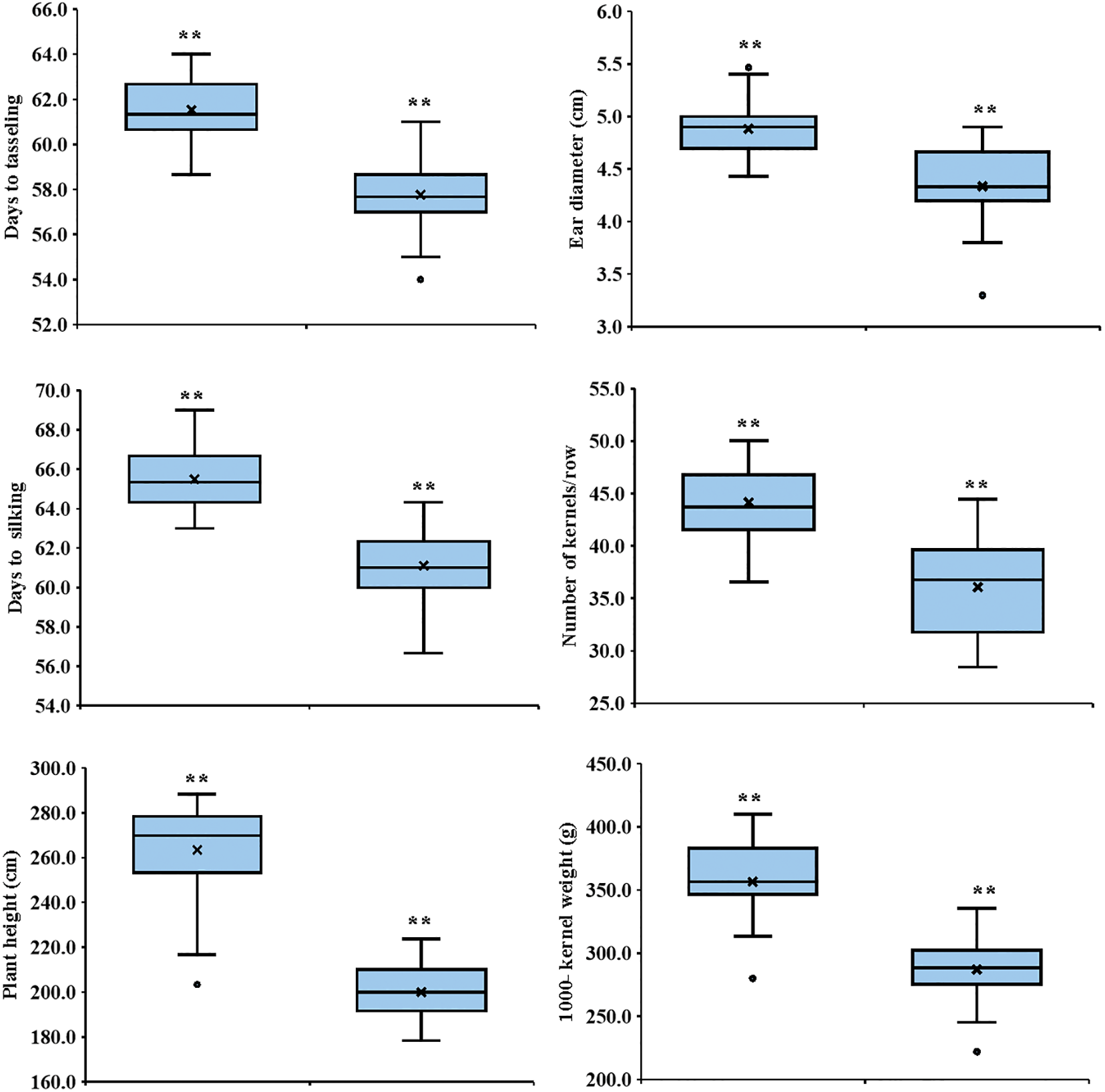
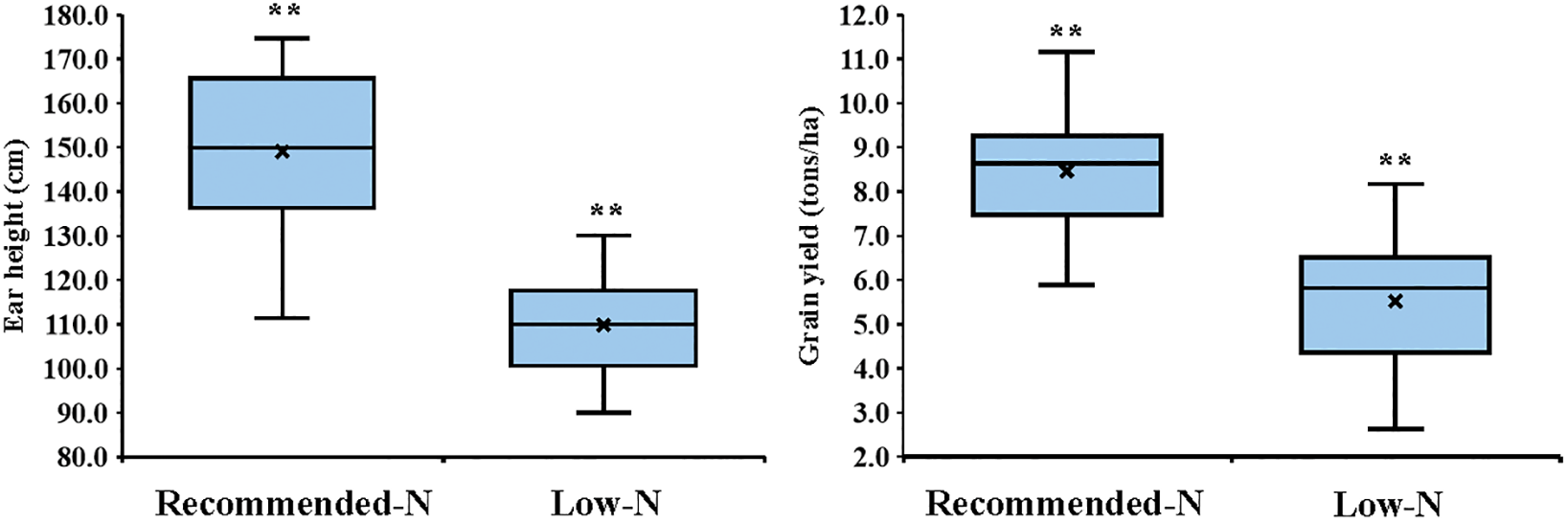
Figure 2: Boxplots displaying the minimum, median, mean, and maximum values for the agronomic traits under study. ** indicates highly significant differences between recommended and low nitrogen levels based on the t-test
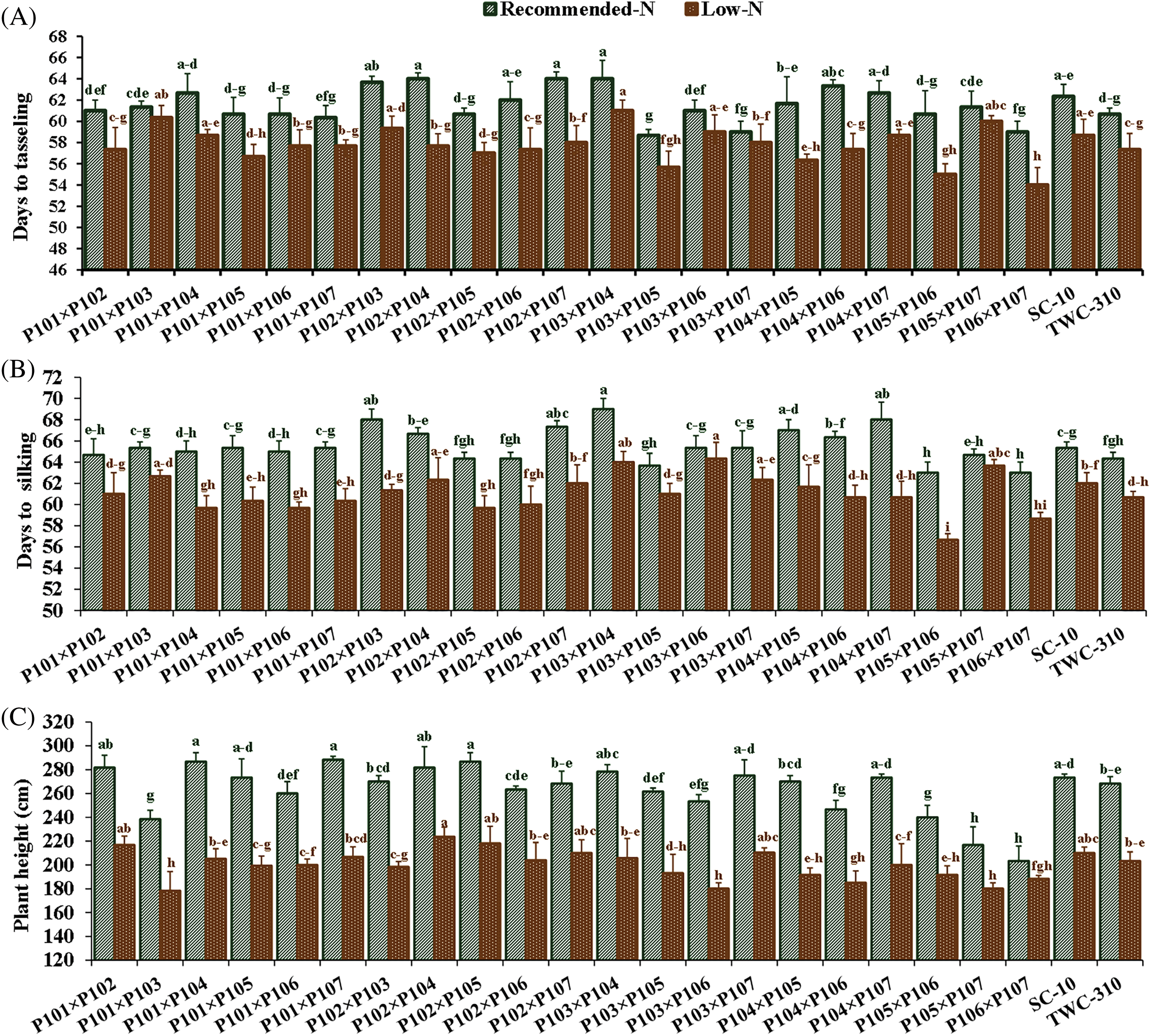
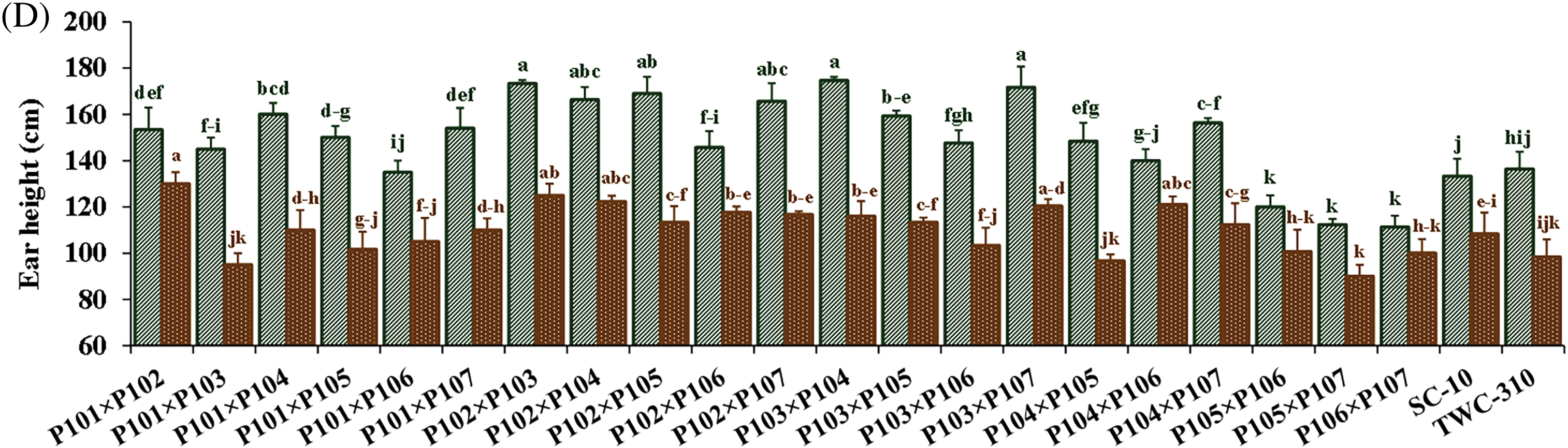
Figure 3: Comparative performance of the assessed twenty-one hybrids and checks hybrids under recommended and low nitrogen levels: Days to tasseling (A), days to silking (B), plant height (C), and ear height (D). The bars above the columns indicate standard deviations (SD). Columns with different letters imply significant differences as determined by LSD with a p-value of less than 0.05
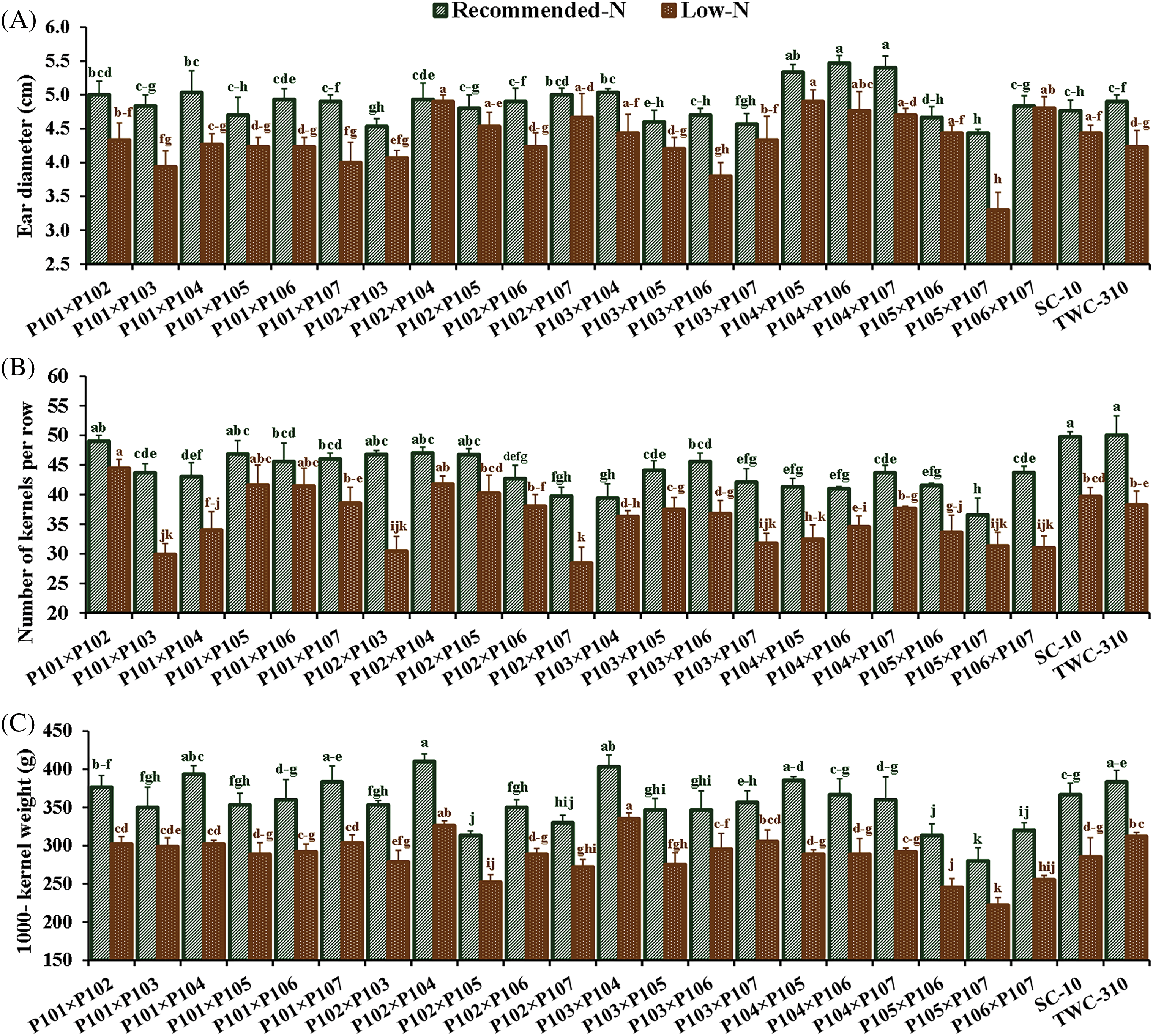
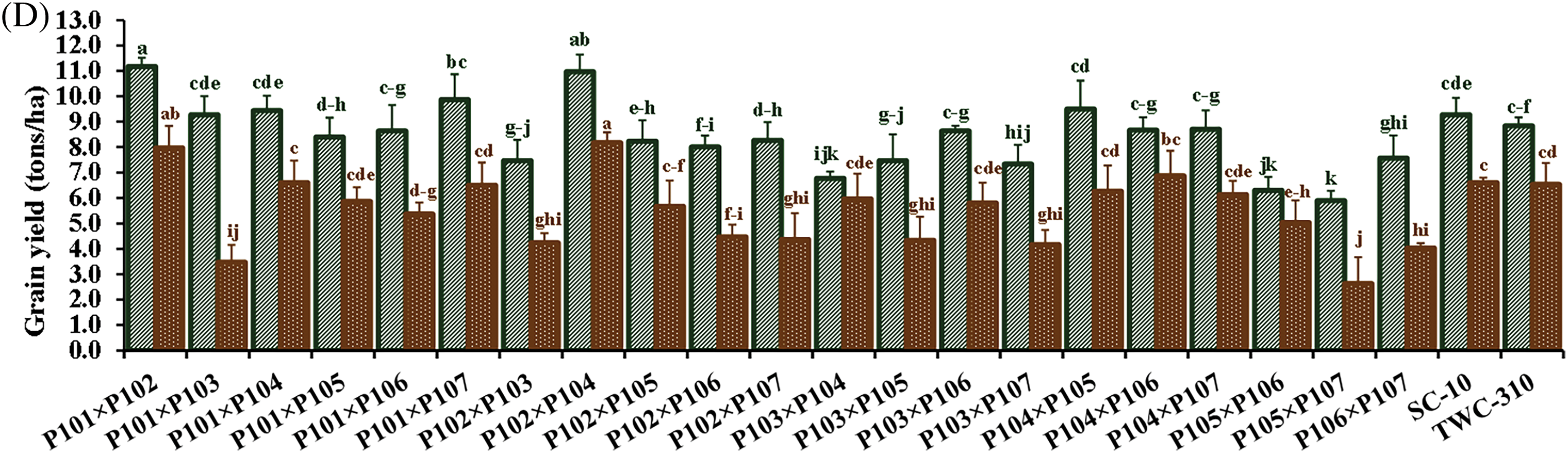
Figure 4: Comparative performance of the assessed twenty-one hybrids and checks hybrids under recommended and low nitrogen levels: Ear diameter (A), number of kernels per row (B), 1000-kernel weight (C) and grain yield (D). The bars above the columns indicate standard deviations (SD). Columns with different letters imply significant differences as determined by LSD with a p-value of less than 0.05
The 21 assessed F1 hybrids and two commercial hybrids (SC-10 and TWC-310) were clustered based on agronomic traits under low nitrogen conditions. These hybrids were divided into five groups, as shown in Fig. 5. Group A, which included two hybrids (P101 × P102 and P102 × P104), recorded the highest agronomic traits. Following group B consisted of seven hybrids (P101 × P107, P104 × P107, P101 × P104, P103 × P104, P104 × P105, P103 × P106, and P104 × P106) characterized by high agronomic performance. Group C contained seven hybrids (SC-10, TWC-310, P101 × P105, P101 × P106, P102 × P105, P102 × P106, and P103 × P105) that showed moderate yield traits. Conversely, Group D included three hybrids (P102 × P107, P102 × P103, and P103 × P107), and Group E included four hybrids (P105 × P106, P106 × P107, P101 × P103, and P105 × P107), both of which encompassed hybrids with the lowest agronomic performance under low nitrogen conditions.
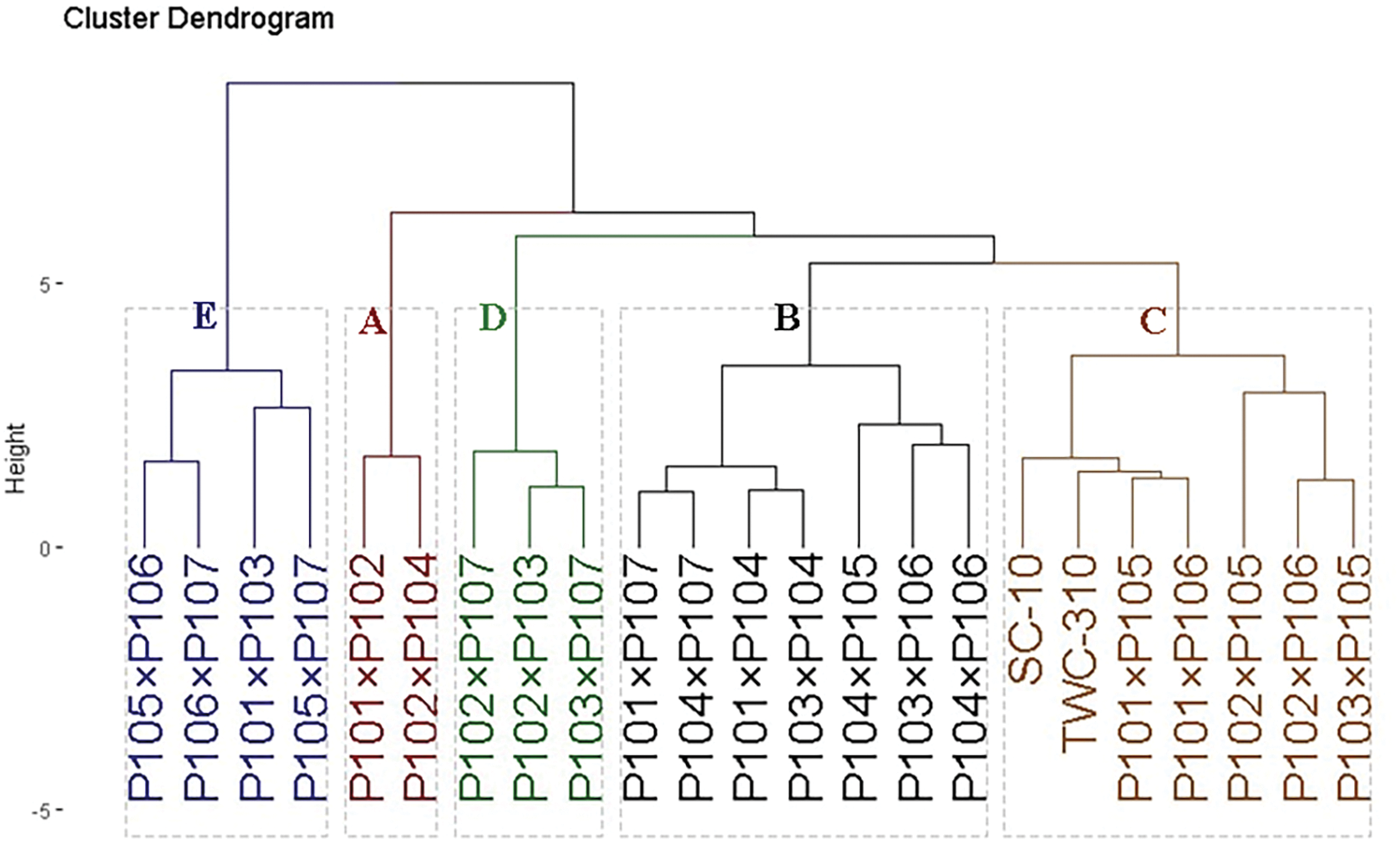
Figure 5: Dendrogram depicting phenotypic distances among 21 hybrids and two commercial hybrids based on agronomic traits
The cross combinations P103 × P105 and P106 × P107 demonstrated the most significant negative heterotic effects for days to tasseling, indicating earlier flowering compared to the commercial hybrids SC-10 under both conditions (Table 2). Likewise, the hybrids P101 × P104, P101 × P106, and P102 × P105 under low N, and P105 × P106 and P106 × P107 under both conditions exhibited the most favorable heterotic effects compared to the commercial hybrids SC-10 for days to silking. For plant height, the crosses P101 × P103, P103 × P106, P104 × P106, P105 × P107, and P106 × P107 exhibited the most favorable heterotic effects, showing shorter plants compared to SC-10 and TWC-310 under both conditions. The most significant negative heterotic effects for ear height, indicating lower ear placement, were observed in P101 × P103, P104 × P105, and P105 × P107 relative to SC-10 under low N conditions. Conversely, the hybrids P104 × P106 and P104 × P107 exhibited the highest positive heterotic effects for ear diameter compared to SC-10 and TWC-310 under normal conditions (Table 3). Additionally, the crosses, P102 × P104, P104 × P105, P104 × P106, and P106 × P107 exhibited the highest positive heterotic effects for ear diameter, surpassing the TWC-310 check hybrid under low N conditions. The best heterotic combinations for number of kernels/row were P101 × P102 and P102 × P104, compared to SC-10 and TWC-310 under low N conditions. The hybrids P102 × P104 and P103 × P104 showed superior positive heterotic effects for thousand kernel weight relative to SC-10 and TWC-310 under both conditions. Additionally, the cross combinations P101 × P102 and P102 × P104 exhibited the highest significant positive heterotic effects for grain yield relative to SC-10 and TWC-310 under both conditions, suggesting these combinations offer superior yield potential due to their advantageous traits.
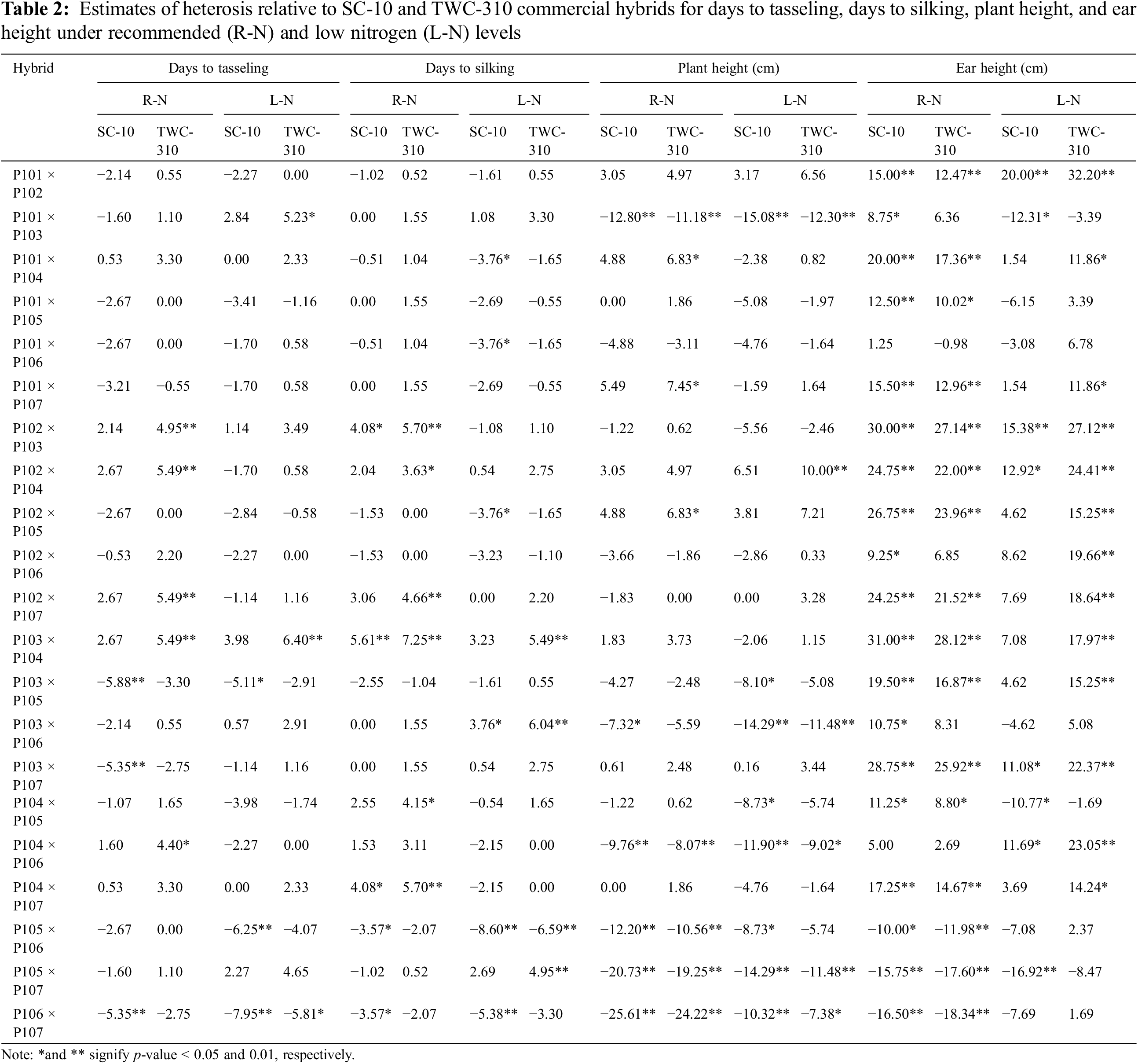
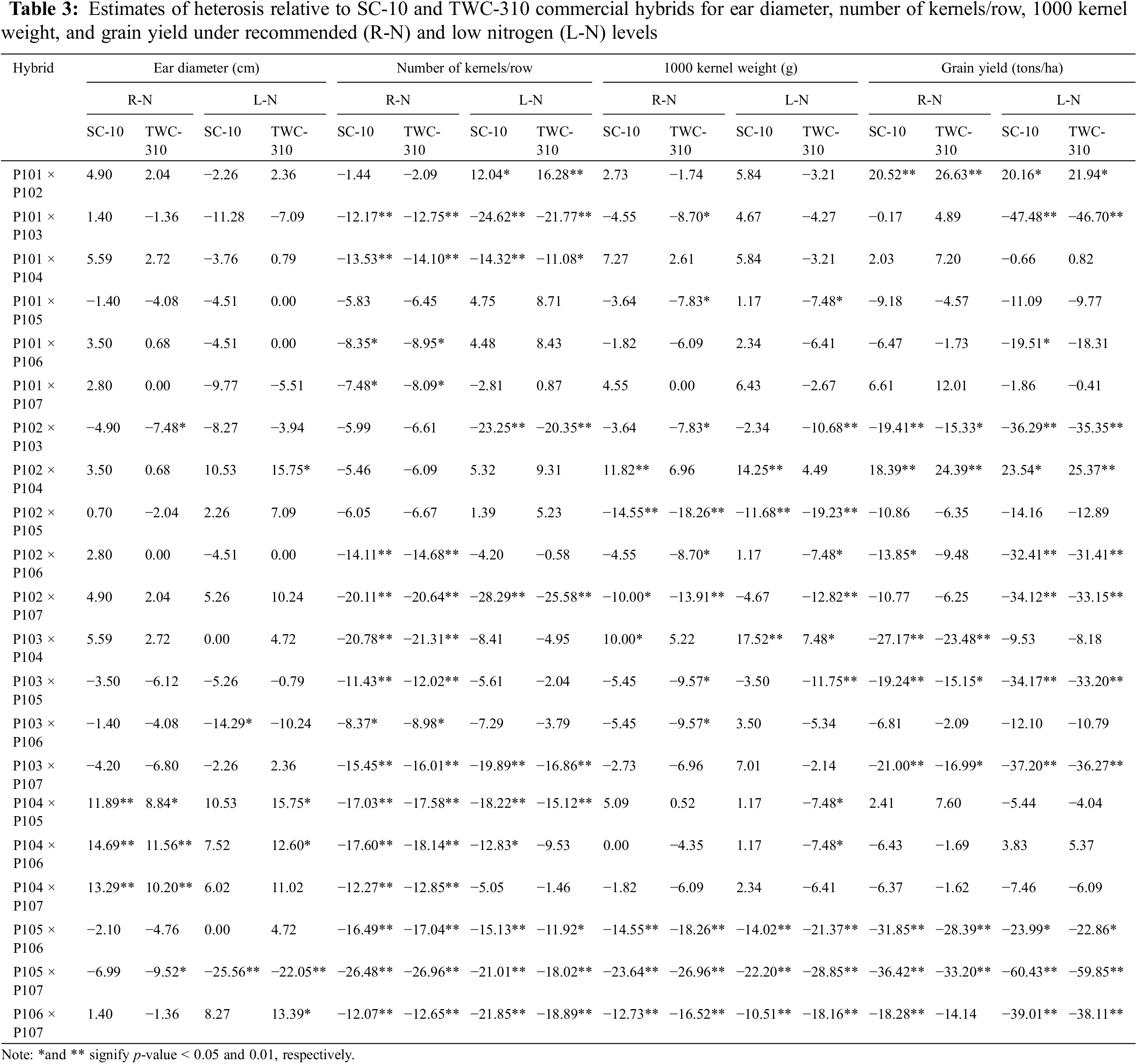
The general combining ability estimated were notably positive and significant for most agronomic traits, except for days to tasseling, days to silking, plant height, and ear height, where negative values are preferable (Table 4). Among the evaluated inbred lines, P105 and P106 exhibited the most significant negative GCA effects for both days to tasseling and days to silking under recommended and low N conditions, indicating their contribution to earlier flowering. Lines P105, P106, and P107 showed the most favorable GCA effects for plant height under recommended conditions and P103, P105, and P106 under low N conditions. The inbreds P105 and P106 emerge as the most effective combiners for ear height under both conditions, attributed to their desirable negative GCA effects. Conversely, P104 demonstrated the highest positive and significant GCA effects for ear length. Lines P101 and P102 had the strongest positive and significant GCA effects for number of kernels per row under both conditions. The superior GCA effects for the thousand kernel weight were attributed to the lines P101 and P104. Under both conditions, the highest GCA effects for grain yield were observed in lines P101, P102, and P104.
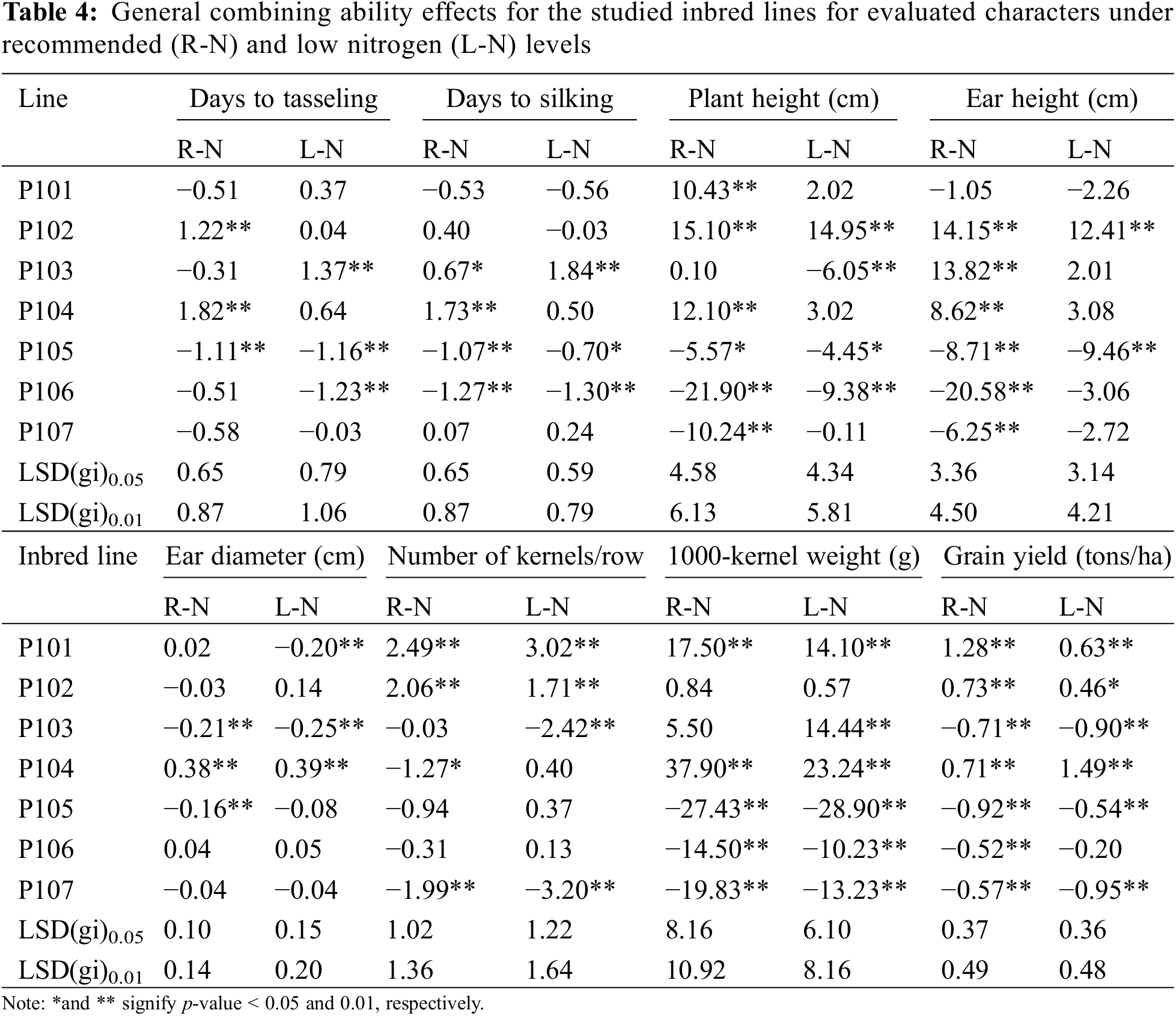
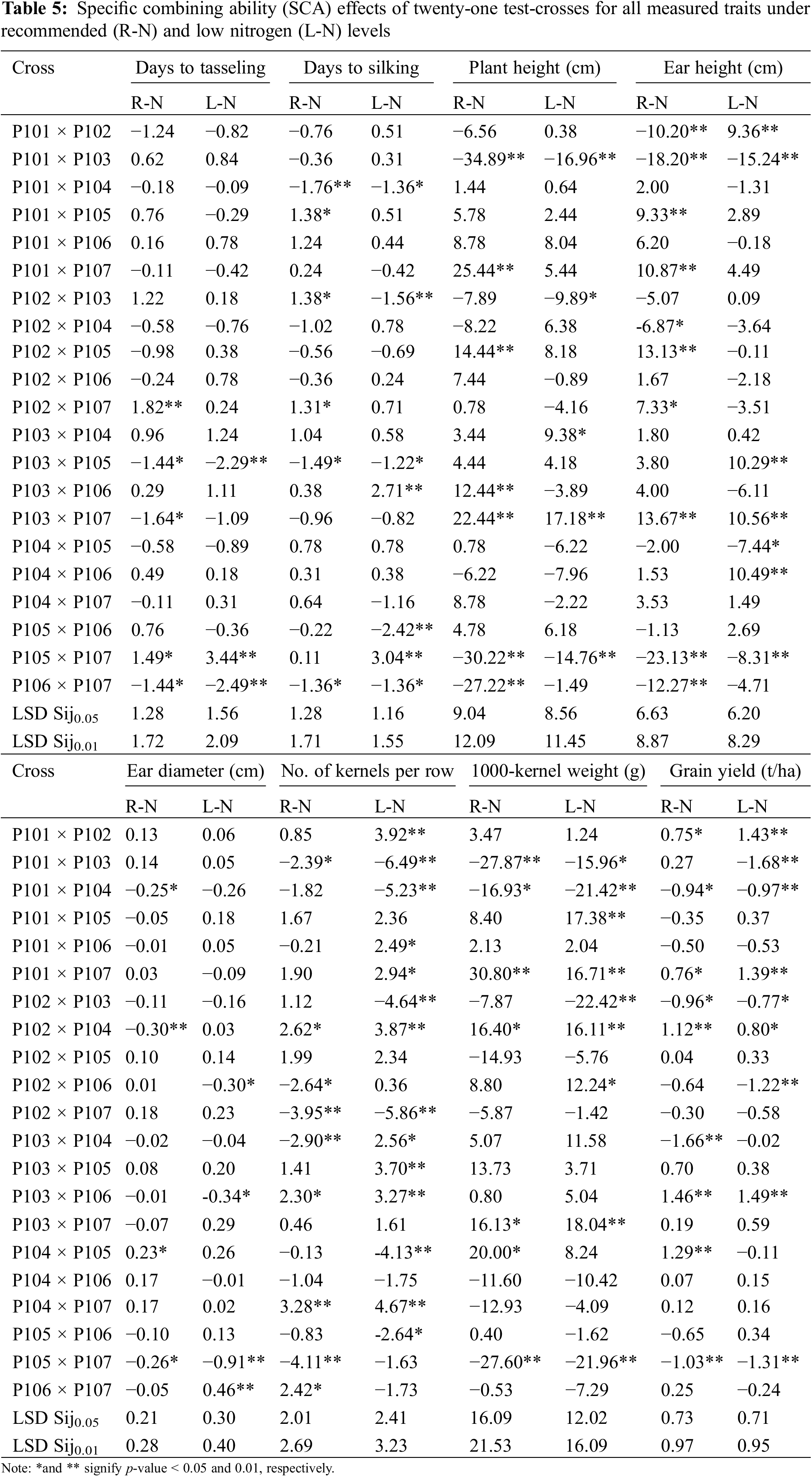
The results presented in Table 5 indicate that the crosses P103 × P105 and P106 × P107 demonstrated the most significant and favorable negative SCA effects for days to silking and days to tasseling under recommended and low N conditions. For ear height and plant height, the crosses P101 × P103 and P105 × P107 displayed the most favorable negative SCA effects under both conditions. In contrast, the highest significant and positive SCA effects for ear diameter were observed in the crosses P104 × P105 under normal conditions and P106 × P107 under low N conditions. Likewise, the advantageous SCA effect for the number of kernels/row was noticed in the hybrids P106 × P107 under recommended conditions, P101 × P102, P101 × P106, P101 × P107, P103 × P104 and P103 × P105 under low N conditions, and P102 × P104, P103 × P106, and P104 × P107 under both conditions. The highest substantial and positive SCA values for thousand kernel weight were recorded in the hybrids P101 × P107, P102 × P104, and P103 × P107 under both conditions. Notably, the hybrids P101 × P102, P101 × P107, P102 × P104, and P103 × P106 demonstrated significantly positive and desirable SCA effects for grain yield under recommended and low N conditions. Although no single hybrid showed optimal SCA effects across all evaluated traits, some hybrids with advantageous effects on grain yield also performed well in one or more related characteristics. For example, the hybrid P101 × P107 showed desirable SCA effects for number of kernels/row, grain yield and thousand-kernel weight. At the same time, the crosses P102 × P104 and P103 × P106 had favorable SCA effects for both grain yield and number of kernels per row under recommended and low N conditions.
3.7 Interrelation among Assessed Hybrids and Studied Traits
The association between the developed maize hybrids and studied agronomic traits was analyzed using principal component (PC) analysis. The first two PCs captured a substantial portion of the variance (59.49% by PC1 and 14.70% by PC2) and were illustrated in the PC biplot (Fig. 6). PC1 exhibited higher variation and efficiently separated the hybrids into those with negative and positive values along this axis. The hybrids on the positive side of PC1 were linked to higher yield traits, notably P102 × P104, P101 × P102, P103 × P104, P103 × P107, and P102 × P107. In contrast, hybrids on the negative side, such as P105 × P107, P101 × P103, P103 × P106, and P106 × P107, demonstrated lower agronomic performance. The closeness of vectors demonstrated a robust positive association among the characters. In particular, grain yield was positively associated with ear height, plant height, 1000-grain weight, and the number of kernels per row. Heatmap and hierarchical clustering based on these traits separated the maize hybrids into diverse clusters (Fig. 7). The hybrids like P102 × P104, P103 × P104, P101 × P102, and P102 × P107 exhibited the highest trait values (represented in blue), whereas P105 × P106 and P106 × P107 displayed the lowest (characterized in red).
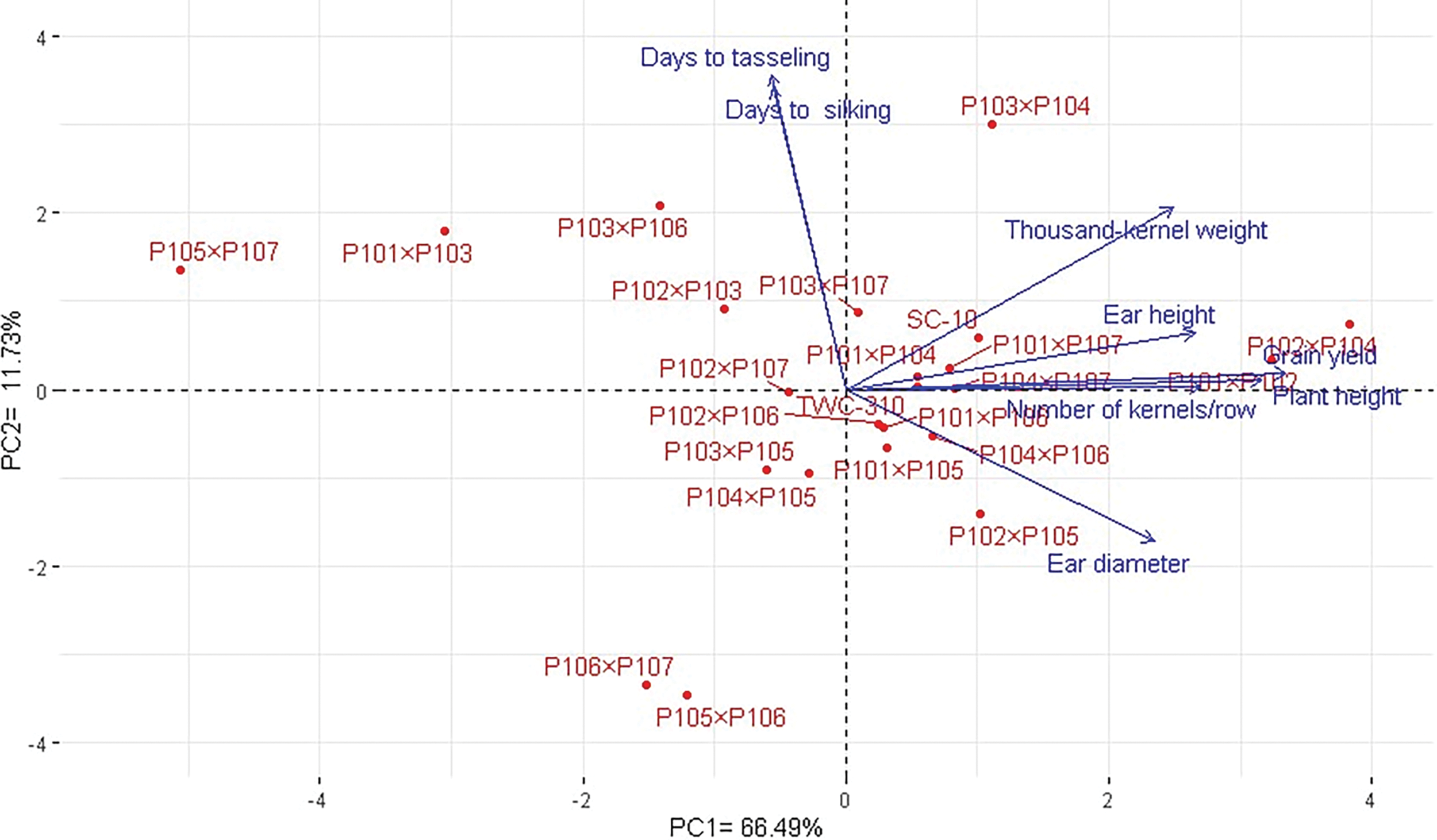
Figure 6: PC-biplot explores the relationship between studied agronomic traits and the developed maize hybrids
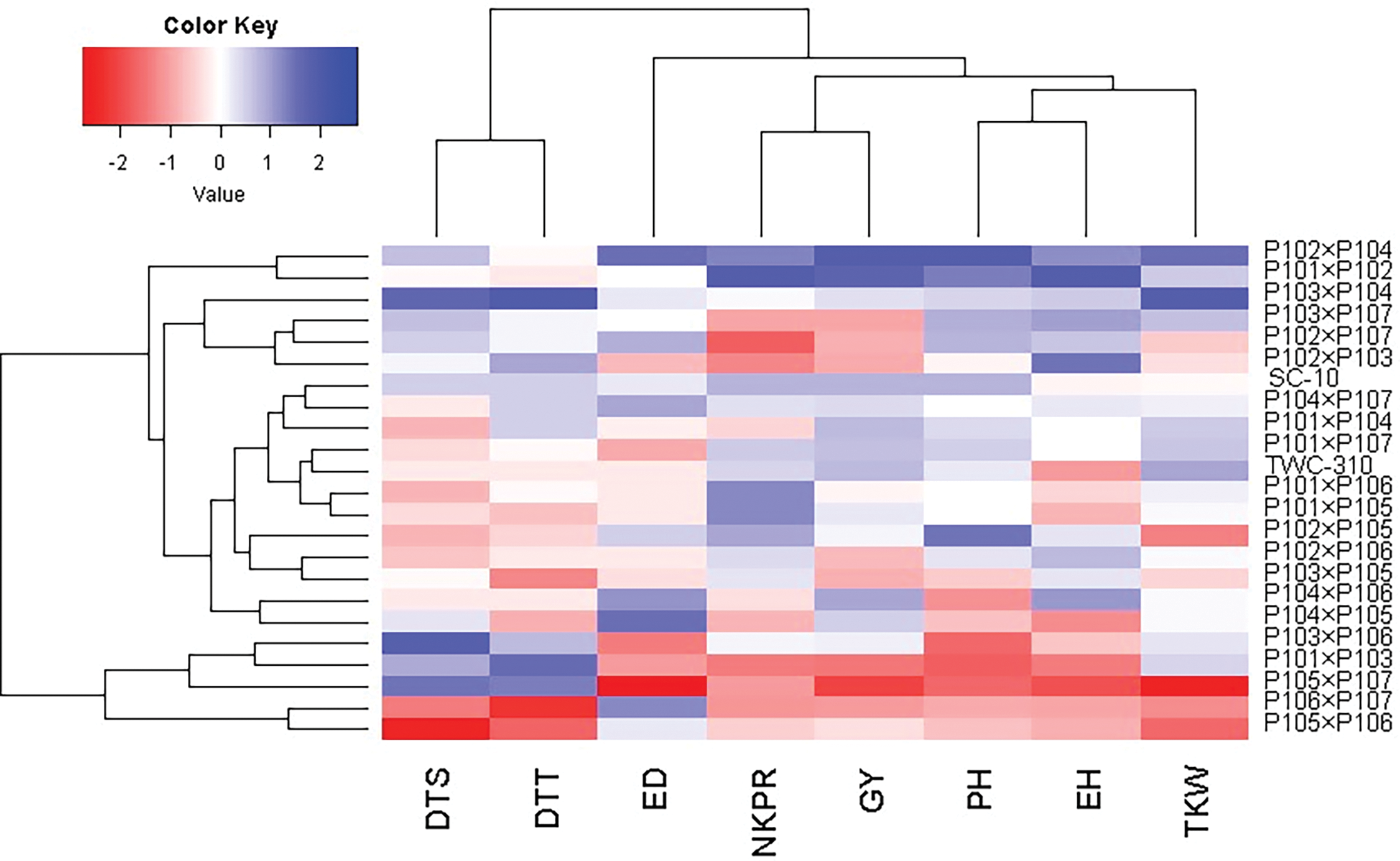
Figure 7: Heatmap and hierarchical clustering analysis to explore the relationships among agronomic attributes in developed maize hybrids and control checks. The traits analyzed included days to silking (DTS), and days to tasseling (DTT), ear diameter (ED), number of kernels/row (NKPR), grain yield (GY), plant height (PH), ear height (EH), and 1000 kernel weight (TKW)
Excessive nitrogen application can result in numerous environmental and ecological problems, such as water pollution, biodiversity loss, and greenhouse gas emissions. Therefore, breeding high-yielding hybrids adapted to low nitrogen conditions is essential to ensure food security, allow cultivation in nitrogen-limited environments, and reduce environmental pollution. Achieving this goal requires a deep understanding of combining ability mechanisms. Genetic variation is the key to developing promising hybrids with improved yield under optimal and low N conditions. Our study highlights significant variations among the evaluated hybrids, demonstrating substantial genetic diversity that supports the selection of superior hybrids under both recommended and low N conditions. This aligns with findings from recent studies, including those by Sedhom et al. [13–15] which detected high genetic variability for several agronomic characters in maize under normal and low N conditions. Developing low N tolerant and high-yielding hybrids principally depends on selecting suitable parental lines [16]. Recognizing diverse lines with beneficial alleles is essential to confirm their effective transmission to progeny [17]. In this context, the favorable GCA effects observed for the inbreds P105 and P106, particularly in relation to tasseling and silking dates, indicate their potential for improving earliness in hybrid maize. Early flowering in maize has essential advantages, such as conserving irrigation water and providing resistance against certain damaging corn borer pests. Shortened time to flowering leaves less time for pest infestation by species such as the African pink borer (Sesamia cretica), pink spotted borer (Pyrausta nubilialis), and spotted borer (Chilo simplex). The inbred lines P105 and P106 were recognized as the most effective general combiner for achieving shorter plant heights and lower ear placement under both nitrogen treatments. This indicates that these two lines have a strong propensity to produce hybrids with reduced plant height, a key trait for developing genotypes more resistant to lodging [18]. Additionally, shorter hybrids might be more efficient at allocating photosynthates towards ear development rather than biomass accumulation [13]. The findings highlighted the inbred line P105 as an effective combiner for achieving early maturity and lower ear placement across low and recommended nitrogen conditions. This indicates its potential for use in breeding early-maturing genotypes with optimal ear placement under both conditions. Improving grain yield and its related traits can be effectively achieved by utilizing the inbreds P101, P102, and P104, which demonstrated high significant and positive GCA effects under low N conditions. This suggests that these inbreds carry valuable alleles that can be passed on to their progeny to produce high-yielding hybrids under nitrogen deficiency conditions. These results align with Okunlola et al. [7] and Luz et al. [19], who observed similar outcomes when using different maize inbreds in nitrogen-stressed environments.
SCA estimates are valuable for identifying promising hybrids. In the present study, most assessed crosses exhibited appropriate and significant SCA effects for at least one attribute. Specifically, crosses P103 × P105 and P106 × P107, which had negative SCA for days to silking and days to tasseling are proper for enhancing earliness in maize under optimal and low N conditions. Additionally, crosses P101 × P102, P101 × P107, P102 × P104, and P103 × P106 showed the strongest SCA for yield traits under both conditions, indicating their potential as high-yielding maize hybrids under varying nitrogen levels. The high SCA resulted from non-additive gene action. Two hybrids, P101 × P102 and P102 × P104, combined desirable SCA with high grain yield under low nitrogen conditions. Overall, these crosses will undergo further evaluation for commercial cultivation.
Understanding trait inheritance patterns is crucial for breeding efforts. This study found that additive and non-additive genetic effects play significant roles in the inheritance of all traits examined, as indicated by substantial GCA and SCA effects. However, the ratio GCA to SCA was less than one for nearly all traits, suggesting that non-additive gene action has a relatively greater influence on the inheritance of these traits under both recommended and low N conditions. Consequently, cross-breeding methods are highly effective for improving these characteristics by leveraging heterosis effects. These results align with those of Akinwale et al. [20] and Badu-Apraku et al. [21]. In contrast to the present findings of this study, Mukaro et al. [22], Habiba et al. [23] and Derera et al. [24] reported that additive gene action plays a predominant role in the inheritance of maize grain yield under normal and low nitrogen conditions.
Exploring the associations between grain yield and its attributes can enhance breeding programs by guiding selection and crosses to improve multiple important characteristics simultaneously [25–27]. The PC-biplot offers an effective means to evaluate the interrelations among the evaluated traits [28–31]. Robust positive associations were found between grain yield and traits such as plant height, number of kernels/row, and 1000-kernel weight under low nitrogen conditions. These findings align with Kamara et al. [17] and Nemati et al. [32], who elucidated a positive relationship between maize grain yield and its attributed traits.
Breeding high-yielding and early maturing maize hybrids under low and recommended nitrogen conditions is crucial for advancing sustainable agriculture and meeting global food demands. The findings exhibited substantial genetic diversity among the assessed maize hybrids, supporting the selection of superior hybrids under both recommended and low N conditions, which is critical for advancing sustainable agriculture. The favorable GCA effects detected for the inbreds P105 and P106, particularly in relation to tasseling and silking dates, highlight their potential for improving earliness in hybrid maize, emphasizing the importance of identifying diverse lines with beneficial alleles for effective transmission to progeny. Several promising hybrids with strong SCA for yield traits were identified under both recommended and low N conditions, particularly the crosses P101 × P102, P101 × P107, P102 × P104, and P103 × P106. Notably, P101 × P102 and P102 × P104 combined desirable SCA with high grain yield, showing potential for commercial cultivation. Strong positive relationships were found between grain yield and traits such as number of kernels/row, plant height, and 1000-kernel weight. Due to their ease of measurement, these traits are helpful for indirect selection to improve grain yield under low nitrogen-stressed environments.
Acknowledgement: The authors are grateful to the Faculty of Agriculture, Kafrelsheikh University, Egypt, for the support provided to conduct this work. The authors also would like to express their gratitude to Princess Nourah bint Abdulrahman University Research Supporting Project Number PNURSP2025R241, Princess Nourah bint Abdulrahman University, Riyadh, Saudi Arabia.
Funding Statement: This project was funded by Princess Nourah bint Abdulrahman University Research Supporting Project Number PNURSP2025R241, Princess Nourah bint Abdulrahman University, Riyadh, Saudi Arabia.
Author Contributions: The authors confirm contribution to the paper as follows: study conception and design: Mohamed M. Kamara, Nora M. Al Aboud, Hameed Alsamadany, Abeer M. Kutby, Imen Ben Abdelmalek, Motrih Al-Mutiry, Diaa Abd El-Moneim; data collection: Mohamed M. Kamara, Nora M. Al Aboud, Diaa Abd El-Moneim; analysis and interpretation of results: Hameed Alsamadany, Abeer M. Kutby, Imen Ben Abdelmalek, Motrih Al-Mutiry; draft manuscript preparation: Hameed Alsamadany, Abeer M. Kutby, Imen Ben Abdelmalek, Motrih Al-Mutiry. All authors reviewed the results and approved the final version of the manuscript.
Availability of Data and Materials: The data are available from the corresponding author upon reasonable request.
Ethics Approval: Not applicable.
Conflicts of Interest: The authors declare no conflicts of interest to report regarding the present study.
References
1. Ahmad U, Hussain M, Ahmad W, Javed J, Arshad Z, Akram Z. Impact of global climate change on maize (Zea maysphysiological responses and modern breeding techniques. Trends Biotechnol Plant Sci. 2024;2:62–77. doi:10.62460/TBPS/2024.020. [Google Scholar] [CrossRef]
2. FAOSTAT. Food and agriculture organization of the united nations. Statistical Database. 2024. Available from: http://www.fao.org/faostat/en/#data. [Accessed 2024]. [Google Scholar]
3. Bakala HS, Devi J, Ankita, Sarao LK, Kaur S. Utilization of wheat and maize waste as biofuel source. In: Agroindustrial waste for green fuel application. Singapore: Springer; 2023. p. 27–66. doi:10.1007/978-981-19-6230-1. [Google Scholar] [CrossRef]
4. Han Z, Hou H, Yao X, Qian X, Zhou M. Substituting partial chemical fertilizers with bio-organic fertilizers to reduce greenhouse gas emissions in water-saving irrigated rice fields. Agronomy. 2024;14:544. doi:10.3390/agronomy14030544. [Google Scholar] [CrossRef]
5. Mohanty LK, Singh N, Raj P, Prakash A, Tiwari AK, Singh V, et al. Nurturing crops, enhancing soil health, and sustaining agricultural prosperity worldwide through agronomy. J Exp Agricul Int. 2024;46:46–67. doi:10.9734/jeai/2024/v46i22308. [Google Scholar] [CrossRef]
6. Rehman Au, Dang T, Qamar S, Ilyas A, Fatema R, Kafle M, et al. Revisiting plant heterosis: from field scale to molecules. Genes. 2021;12:1688. doi:10.3390/genes12111688. [Google Scholar] [CrossRef]
7. Okunlola G, Badu-Apraku B, Ariyo O, Ayo-Vaughan M. The combining ability of extra-early maturing quality protein maize (Zea mays) inbred lines and the performance of their hybrids in Striga-infested and low-nitrogen environments. Front Sustain Food Syst. 2023;7:1238874. doi:10.3389/fsufs.2023.1238874. [Google Scholar] [CrossRef]
8. Amzeri A, Suhartono FS, Pawana G, Sukma K. Combining ability analysis in maize diallel hybrid. SABRAO J Breed Genet. 2024;56:476–92. doi:10.54910/sabrao2024.56.2.3. [Google Scholar] [CrossRef]
9. Arora A, Das AK, Dixit S, Y. KR, Singh S, Sekhar J, et al. Genetic diversity analysis and heterotic grouping of Indian white maize inbred lines using combining ability and SNP markers. Crop Sci. 2024;64:726–37. doi:10.1002/csc2.v64.2. [Google Scholar] [CrossRef]
10. Heiba H, Mahgoub E, Mahmoud A, Ibrahim M, Mansour E. Combining ability and gene action controlling chocolate spot resistance and yield traits in faba bean (Vicia faba L.). J Agricul Sci. 2023;29:77–88. [Google Scholar]
11. Akinwale R, Badu-Apraku B, Fakorede M, Vroh-Bi I. Heterotic grouping of tropical early-maturing maize inbred lines based on combining ability in Striga-infested and Striga-free environments and the use of SSR markers for genotyping. Field Crops Res. 2014;156:48–62. doi:10.1016/j.fcr.2013.10.015. [Google Scholar] [CrossRef]
12. Griffing B. Concept of general and specific combining ability in relation to diallel crossing systems. Aust J Biol Sci. 1956;9(4):463–93. doi:10.1071/BI9560463. [Google Scholar] [CrossRef]
13. Sedhom YS, Rabie HA, Awaad HA, Alomran MM, ALshamrani SM, Mansour E, et al. Genetic potential of newly developed maize hybrids under different water-availability conditions in an arid environment. Life. 2024;14(4):453. doi:10.3390/life14040453. [Google Scholar] [CrossRef]
14. Kamara MM, Mansour E, Khalaf AE, Eid MA, Hassanin AA, Abdelghany AM, et al. Molecular diversity and combining ability in newly developed maize inbred lines under low-nitrogen conditions. Life. 2024;14(5):641. doi:10.3390/life14050641. [Google Scholar] [CrossRef]
15. Engida BT, Wegary D, Keno T, Mekonnen TW. Combining ability and genetic distance analysis of mid altitude sub-humid agroecology adapted maize inbred lines for high grain yield. Heliyon. 2024;10(11):e32267. doi:10.1016/j.heliyon.2024.e32267. [Google Scholar] [CrossRef]
16. Adu GB, Badu-Apraku B, Akromah R. Strategies for selecting early maturing maize inbred lines for hybrid production under low soil nitrogen and Striga infestation. Agronomy. 2021;11(7):1309. doi:10.3390/agronomy11071309. [Google Scholar] [CrossRef]
17. Kamara MM, Ghazy NA, Mansour E, Elsharkawy MM, Kheir AM, Ibrahim KM. Molecular genetic diversity and line× tester analysis for resistance to late wilt disease and grain yield in maize. Agronomy. 2021;11(5):898. doi:10.3390/agronomy11050898. [Google Scholar] [CrossRef]
18. Omar M, Rabie HA, Mowafi SA, Othman HT, El-Moneim DA, Alharbi K, et al. Multivariate analysis of agronomic traits in newly developed maize hybrids grown under different agro-environments. Plants. 2022;11(9):1187. doi:10.3390/plants11091187. [Google Scholar] [CrossRef]
19. Luz LS, Reis HMD, De Leon da Costa NMP, Carvalho FR, Caixeta DG, DeLima RO. Combining ability for agronomic traits among commercial maize hybrids under low and high nitrogen inputs targeting the development of breeding populations. PLoS One. 2024;19(10):e0309296. doi:10.1371/journal.pone.0309296. [Google Scholar] [CrossRef]
20. Akinwale RO, Eze CE, Traore D, Menkir A. Detection of non-additive gene action within elite maize populations evaluated in contrasting environments under rainforest ecology in Nigeria. Crop Breed, Genet Genomics. 2021;3:1–24. [Google Scholar]
21. Badu-Apraku B, Oyekunle M, Akinwale RO, Aderounmu M. Combining ability and genetic diversity of extra-early white maize inbreds under stress and nonstress environments. Crop Sci. 2013;53(1):9–26. doi:10.2135/cropsci2012.06.0381. [Google Scholar] [CrossRef]
22. Mukaro R, Kamutando CN, Magorokosho C, Mutari B, Zaidi PH, Kutywayo D, et al. Genetic potential of tropically adapted exotic maize (Zea mays L.) heat-tolerant donor lines in sub-tropical breeding programs. Agronomy. 2023;13:2050. doi:10.3390/agronomy13082050. [Google Scholar] [CrossRef]
23. Habiba RMM, El-Diasty MZ, Aly RSH. Combining abilities and genetic parameters for grain yield and some agronomic traits in maize (Zea mays L.). Beni-Suef Univ J Basic Appl Sci. 2022;11:108. doi:10.1186/s43088-022-00289-x. [Google Scholar] [CrossRef]
24. Derera J, Tongoona P, Vivek BS, Laing MD. Gene action controlling grain yield and secondary traits in southern African maize hybrids under drought and non-drought environments. Euphytica. 2008;162:411–22. doi:10.1007/s10681-007-9582-4. [Google Scholar] [CrossRef]
25. Kamara MM, Rehan M, Mohamed AM, El Mantawy RF, Kheir AM, Abd El-Moneim D, et al. Genetic potential and inheritance patterns of physiological, agronomic and quality traits in bread wheat under normal and water deficit conditions. Plants. 2022;11:952. doi:10.3390/plants11070952. [Google Scholar] [CrossRef]
26. ElShamey EA, Sakran RM, ElSayed MA, Aloufi S, Alharthi B, Alqurashi M, et al. Heterosis and combining ability for floral and yield characters in rice using cytoplasmic male sterility system. Saudi J Biol Sci. 2022;29:3727–38. doi:10.1016/j.sjbs.2022.03.010. [Google Scholar] [CrossRef]
27. Sakran RM, Ghazy MI, Rehan M, Alsohim AS, Mansour E. Molecular genetic diversity and combining ability for some physiological and agronomic traits in rice under well-watered and water-deficit conditions. Plants. 2022;11(5):702. doi:10.3390/plants11050702. [Google Scholar] [CrossRef]
28. Habibullah M, Sarkar S, Islam MM, Ahmed KU, Rahman MZ, Awad MF, et al. Assessing the response of diverse sesame genotypes to waterlogging durations at different plant growth stages. Plants. 2021;10(11):2294. doi:10.3390/plants10112294. [Google Scholar] [CrossRef]
29. Mannan MA, Tithi MA, Islam MR, Al Mamun MA, Mia S, Rahman MZ, et al. Soil and foliar applications of zinc sulfate and iron sulfate alleviate the destructive impacts of drought stress in wheat. Cereal Res Commun. 2022;50(4):1279–89. doi:10.1007/s42976-022-00262-5. [Google Scholar] [CrossRef]
30. Swailam MA, Mowafy SAE, El-Naggar NZA, Mansour E. Agronomic responses of diverse bread wheat genotypes to phosphorus levels and nitrogen forms in a semiarid environment. SABRAO J Breed Genet. 2021;53(4):592–608. doi:10.54910/sabrao2021.53.4.4. [Google Scholar] [CrossRef]
31. Mansour E, Moustafa ES, Abdul-Hamid MI, Ash-shormillesy SM, Merwad ARM, Wafa HA, et al. Field responses of barley genotypes across a salinity gradient in an arid Mediterranean environment. Agric Water Manag. 2021;258(8):107206. doi:10.1016/j.agwat.2021.107206. [Google Scholar] [CrossRef]
32. Nemati A, Sedghi M, Sharifi RS, Seiedi MN. Investigation of correlation between traits and path analysis of corn (Zea mays L.) grain yield at the climate of Ardabil region (Northwest Iran). Notulae Botanicae Horti Agrobotanici Cluj-Napoca. 2009;37:194–8. [Google Scholar]
Appendix A
Cite This Article
 Copyright © 2025 The Author(s). Published by Tech Science Press.
Copyright © 2025 The Author(s). Published by Tech Science Press.This work is licensed under a Creative Commons Attribution 4.0 International License , which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
 Downloads
Downloads
 Citation Tools
Citation Tools