
 Submit a Paper
Submit a Paper Propose a Special lssue
Propose a Special lssue Open Access
Open Access
REVIEW
Crosstalk between Extracellular Vesicles and the Tumor Microenvironment: Mechanistic Insights and Therapeutic Opportunities
1 State Key Laboratory of Mechanism and Quality of Chinese Medicine, Institute of Chinese Medical Sciences, University of Macau, Macao SAR, China
2 School of Chinese Medicine, Li Ka Shing Faculty of Medicine, The University of Hong Kong, Hong Kong SAR, China
* Corresponding Author: Xiaowen Mao. Email:
(This article belongs to the Special Issue: Targeting the Tumor Microenvironment: Emerging Insights into Cancer Progression and Therapeutics)
Oncology Research 2026, 34(8), 7 https://doi.org/10.32604/or.2026.079562
Received 23 January 2026; Accepted 13 April 2026; Issue published 16 July 2026
 View Full Text
View Full Text Download PDF
Download PDFAbstract
Extracellular vesicles (EVs) are actively secreted, membrane-enclosed nanoparticles that serve as pivotal mediators of intercellular communication. They function as key mediators of intercellular communication by transporting diverse biomolecules, including proteins, nucleic acids, and metabolites. Within the tumor microenvironment, EVs drive complex cellular crosstalk and critically regulate tumor progression by remodeling the extracellular matrix, conferring drug resistance, and reprogramming immune responses. Given their natural biocompatibility, tissue tropism, and ability to cross biological barriers, EVs have emerged as promising platforms for immunotherapy, tumor vaccines and targeted drug delivery system. Moreover, the rapid expansion of EV-based clinical trials highlights their promise in precision medicine. Concurrently, the limitations associated with EV-based therapeutic strategies are critically evaluated to inform future development. This review has also detailed the importance of single-vesicle analysis, which represents a rapidly evolving frontier in EV science. Aiming to bridge the gap between mechanistic understanding and clinical practice, this review delineates the pivotal roles of EV-mediated communication in reshaping the dynamic homeostasis of the tumor microenvironment. Moreover, we provide a critical analysis of current EV-based therapeutic pipelines and their translational challenges, offering key perspectives and theoretical support for the future of precision medicine.Graphic Abstract
Keywords
Intercellular signaling mechanisms are essential for homeostatic maintenance and facilitate rapid cellular adaptation to dynamic environmental changes. Classically, this communication is mediated through direct cell-to-cell contact and the exchange of bioactive molecules [1]. Recent studies have demonstrated that extracellular vesicles (EVs) act as pivotal mediators of intercellular communication, enabling both short- and long-range cellular communication [2].
EVs are lipid bilayer-enclosed nanoparticles released by cells that lack self-replication capacity. Historically, EVs have been named according to various criteria such as size, biogenetic pathway, and cellular origin. In line with the 2023 Minimal Information for Studies of Extracellular Vesicles (MISEV) recommendations, EVs can be broadly categorized by diameter into small EVs (<200 nm) and large EVs (>200 nm). Nevertheless, in biological research, classification based on EVs biogenesis remains more prevalent. Exosomes, typically under 200 nm, derive from the endosomal system [3]. Ectosomes (including microvesicles and microparticles) are generated by direct outward budding of the plasma membrane and exhibit a wider size distribution [3]. Emerging research has identified additional EV populations, including migrasomes [4] and blebbisomes [5], further expanding EV classification.
Common protein markers used to characterize EVs include transmembrane proteins such as tetraspanins proteins (cluster of differentiation 9 [CD9], CD63, CD81) and flotillin-1/2, as well as biomarkers associated with EV biogenesis, including Tumor susceptibility gene 101 (TSG101), apoptosis-linked gene 2-interacting protein X (ALIX), annexin A1, and solute carrier family 3 member 2 (SLC3A2) [3,6,7]. It should be noted that no single biomarker currently exists that adequately captures the heterogeneity of the EV population or can reliably distinguish between EV subtypes.
Under normal physiological conditions, EVs function as key mediators of intercellular communication, facilitating the exchange of bioactive proteins, lipids, and nucleic acids between donor and recipient cells [8]. The composition of EV cargo is dynamic and influenced by the type and state of the donor cells, as well as by changes in the external environment [9]. Once released, EVs can travel via bodily fluids over short or long distances and are taken up by recipient cells through mechanisms such as membrane fusion, receptor-mediated uptake, or endocytosis, thereby enabling the transfer of EV cargo and modulation of recipient cell function [10].
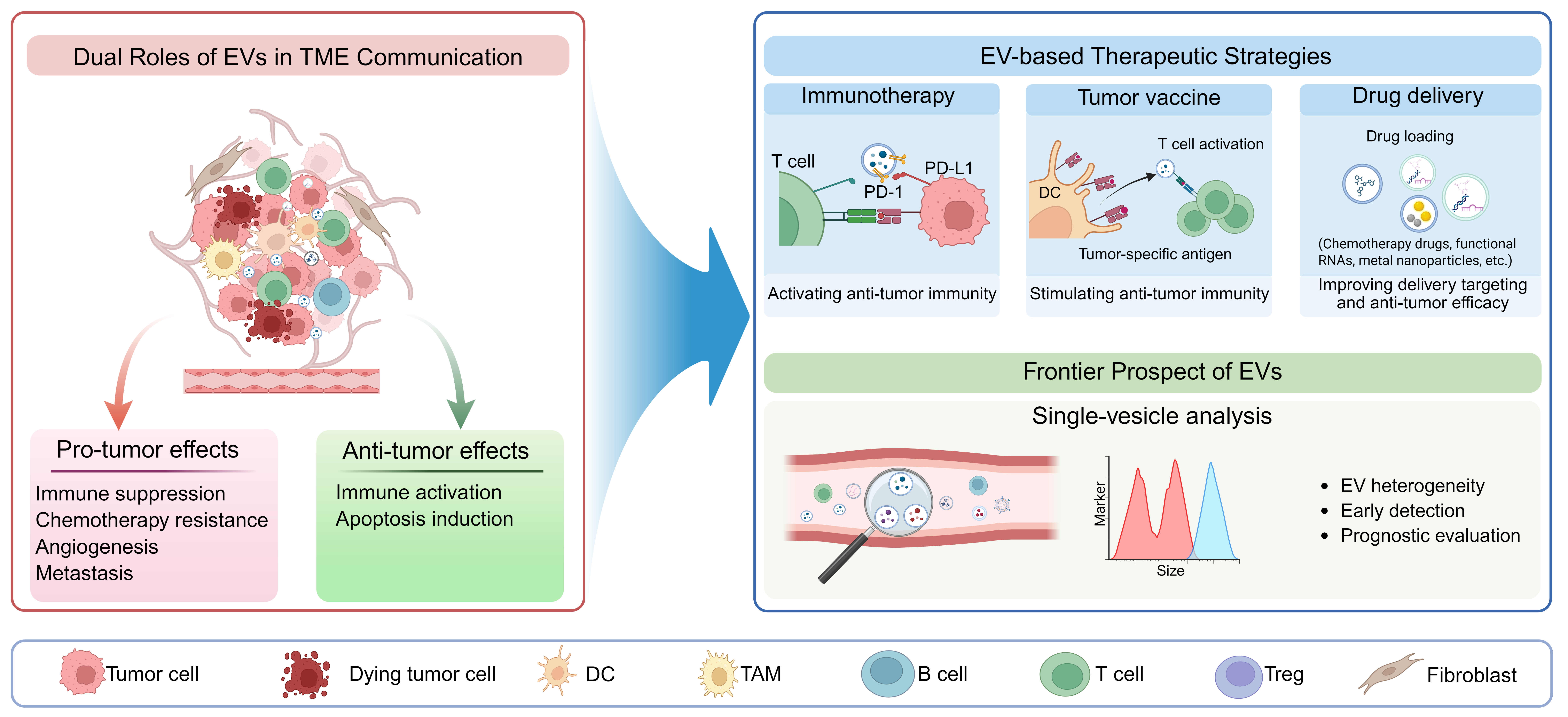
The hypermetabolic state of malignant cells and perturbations of the tumor microenvironment (TME) drive abundant EV release into circulation [11,12]. Tumor cell-derived EVs actively remodel the extracellular matrix (ECM) to establish an environment suitable for tumor cell survival and proliferation [13]. Additionally, EVs within the TME facilitate tumor progression by potentiating tumor angiogenesis [14], participating in pre-metastatic niche formation [15], mediating immune evasion [16] and conferring chemoresistance via transfer of EV cargo [17]. Current efforts are increasingly focused on unraveling the complex interactions between EVs and the TME, alongside the development of novel therapeutic strategies aimed at disrupting EV-mediated intercellular crosstalk in cancer progression. Our review synthesizes emerging evidence on the dynamic interplay between EVs and the TME, elucidating key molecular mechanisms underlying their pro-tumorigenic interactions. Furthermore, we discuss the emerging therapeutic strategies designed to disrupt EV-TME communication for clinical benefit (Fig. 1).

Figure 1: Current Landscape and Future Frontiers of Extracellular Vesicles-Based Therapies in Cancer. Extracellular vesicles (EVs) mediate complex intercellular communication within the tumor microenvironment (TME) and exert binary effects on the tumor progression. On the one hand, EVs promote tumor progression through immunosuppression, chemoresistance, angiogenesis, and metastasis. On the other hand, they also manifest anti-tumor effects via immune activation and the induction of apoptosis. Based on these biological mechanisms, diverse novel therapeutic approaches are being explored, categorizable into immunotherapy, which modulates the TME immune response and stimulates anti-tumor immunity; tumor vaccines, that activate anti-tumor immunity by delivering tumor-specific antigens; and drug delivery, leveraging EVs for cargo loading (e.g., small molecules, siRNA) and targeted delivery to tumor cells. Furthermore, advancements in single-EV analysis techniques facilitate the dissection of EV heterogeneity and hold immense potential for applications in early tumor diagnosis and prognosis assessment. DC: Dendritic Cell; TAM: Tumor-Associated Macrophage.
2 EV-Driven Crosstalk among TME Cells
TME represents a complex, heterogeneous ecosystem characterized by non-cellular structural elements and a sophisticated cellular landscape, including phenotypically distinct tumor cells, immune cells (e.g., dendritic cells, lymphocytes, and macrophages), stromal cells, endothelial cells, and cancer-associated fibroblasts (CAFs) [18]. These cellular constituents actively secrete EVs that mediate intercellular crosstalk, collectively shaping the dynamic TME landscape and driving tumor progression [19].
2.1 Effects of EVs in Tumor Cell
Accumulating evidence identifies EVs as critical mediators of tumor progression and metastatic dissemination. Specifically, EVs participate in multiple steps of the metastatic cascade, either by directly conferring invasive properties to cancer cells or by delivering matrix metalloproteinases (MMPs) that degrade the ECM, thereby facilitating tumor cell dissemination [20,21]. Zhang et al. reported that therapy-induced senescence in colorectal cancer cells drives tumor progression through senescence-associated secretory phenotype-mediated EV secretion. Specifically, senescent cells produce serpin family E member 1-enriched EVs that, following recipient cell uptake, orchestrate NF-κB activation via p65 binding and nuclear translocation, establishing a pro-tumorigenic feedback loop [22].
MicroRNA (miRNA) represent a primary class of functional cargoes encapsulated within EVs, serving as pivotal regulators of post-transcriptional gene expression upon uptake by recipient cells [23]. The study by Li and colleagues revealed that EV-mediated intercellular communication from M2 tumor-associated macrophages (TAMs) to Non-small cell lung cancer (NSCLC) cells involves selective enrichment of oncogenic miRNAs (miR-155/miR-196a-5p), which significantly enhance metastatic potential both in vitro and in vivo [24].
EVs have been established as critical mediators of therapeutic resistance in cancer [25]. EVs contribute to chemotherapy resistance via several key mechanisms. These include the horizontal transfer of functional cargo, such as drug efflux pumps and anti-apoptotic proteins, into recipient cells. This process reduces intracellular drug accumulation and attenuates apoptosis, thereby promoting tumor cell survival under therapeutic pressure. Chemotherapy-induced EVs (chemo-EVs) undergo molecular remodeling, carrying elevated levels of pro-tumorigenic factors that critically influence tumor behavior [26]. Chemotherapeutic agents further enhance EV secretion from tumor cells [27]. Specifically, Kreger et al. demonstrated that that paclitaxel treatment triggers the release of survivin-enriched EVs from breast cancer cells, which promotes cancer cell survival and confers chemoresistance to recipient cells [28].
Furthermore, EVs induce transcriptional reprogramming within tumor cells by delivering oncogenic miRNAs and lncRNAs, activating pro-survival signaling pathways and leading to chemotherapy resistance. The immunosuppressive TAM-EV miRNA cargo plays a pivotal role in therapeutic resistance. Specifically, Binenbaum et al. demonstrated that pancreatic ductal adenocarcinoma (PDAC)-bearing mice treated with macrophage-derived EVs developed resistance to gemcitabine, the first-line therapeutic agent for PDAC. This resistance was mediated by TAM-secreted miR-365, which impairs gemcitabine activity via two distinct pathways. Mechanistically, miR-365 involves the expansion of the triphospho-nucleotide pool, which exerts competitive inhibition on gemcitabine, alongside the induction of cytidine deaminase-mediated drug inactivation [29].
In summary, EVs mediate the transfer of diverse oncogenic cargo within the TME, driving metastatic dissemination and chemoresistance in recipient cancer cells. Therefore, targeting EV-mediated intercellular communication networks may offer a theoretical framework for developing novel strategies to suppress tumor progression and overcome clinical resistance.
2.2 Crosstalk of EVs and Endothelial Cells in the TME
EVs transport diverse protein and nucleic acid cargoes that actively regulate endothelial cells within the TME, promoting angiogenesis within tumors, reducing cell permeability, and facilitating pre-metastatic niche formation [30]. Specifically, EVs from senescent fibroblasts promote tumor angiogenesis and endothelial sprouting through a unique molecular signature marked by CD9 downregulation and ANGPTL2 enrichment [31]. Similarly, gastric cancer-derived EVs (GC-EVs) deliver angiopoietin-2 (ANG2) to activate the Phosphoinositide 3-kinase/protein kinase B (PI3K/AKT) pathway in endothelial cells, thereby stimulating angiogenesis [32]. In metastatic breast cancer, EV-encapsulated miR-105 disrupts endothelial barrier integrity by downregulating tight junction proteins, facilitating hematogenous dissemination. Furthermore, tumor EV-derived non-coding RNAs, including miR-92a-3p [33], miR-183-5p [34], and the long non-coding RNA UBE2CP3 [35], mediate intercellular crosstalk between tumor cells and endothelial cells. S100A16 encapsulated in the endothelial cell-derived EVs (EC-EVs) promotes resistance to apoptosis and facilitates brain tissue colonization by small cell lung cancer cells [36]. These examples underscore the cargo-dependent nature of EC-EV mediated intercellular communication in cancer progression.
Elucidating EV-endothelial cell interactions may help enhance the efficacy of anti-angiogenic drugs and establish a theoretical basis for the development of highly effective targeted drugs.
2.3 Dual Regulation of EVs on Immune Cells in the TME
Tumor-associated macrophages (TAMs) constitute a pivotal cellular component in the TME and are often thought to be associated with poor prognosis [37]. For decades, macrophages have been conventionally classified into two main phenotypes: the pro-inflammatory M1 type and the immunosuppressive M2 type. M1 exert anti-tumor effects and secret pro-inflammatory cytokine, whereas M2 promote tumor progression [38]. The relative proportion of M1 and M2 in the TME serves as a determinant of tumor progression, with a lower M1/M2 ratio typically associated with advanced disease stages and therapy resistance. However, this M1/M2 classification is now considered oversimplified, as it ignores the dynamic heterogeneity of TAMs and the complexity of the TME [39]. Enabled by the rapid advancement of single-cell technologies, TAMs are now subdivided into more refined subpopulations based on distinct functional states and gene signatures. These include interferon-primed TAMs, immune regulatory TAMs, inflammatory cytokine-enriched TAMs, lipid-associated TAMs, pro-angiogenic TAMs, tissue-resident macrophage-like TAMs, and glycolytic TAMs [37,40].
Likewise, other immune cell populations play critical roles within the TME. As innate immune effectors, natural killer (NK) cells eliminate tumor cells through the release of perforin and granzymes upon activation, constituting an important component of cancer immunotherapy [41]. Dendritic cells (DCs), as professional antigen-presenting cells, recognize and process tumor-associated antigens to initiate adaptive immune responses [42]. Among adaptive immune cells, T cells are central to antitumor immunity: helper T cells and cytotoxic T cells respectively coordinate immune responses and directly kill tumor cells, whereas regulatory T cells (Tregs) suppress immune activity and contribute to tumor immune evasion [43].
EVs serve as pivotal mediators of communication between immune cells—such as macrophages, NK cells, T cells, and B cells—and tumor cells in the TME. Through the transfer of immunosuppressive or immunostimulatory molecules, EVs dynamically reprogram the local immune microenvironment.
During tumor initiation, tumor-derived extracellular vesicles (TDEVs) contribute to the formation of a microenvironment centered around M1 macrophages [30]. TDEVs contain cargoes, including miR-3342 [44] and miR-125b-243 [45], which can exert anti-tumor effects by directly polarizing macrophages to M1 or reprogramming M2 macrophages to M1. DC-derived EVs carrying TNF-α activate natural killer (NK) cells via TNF receptor binding, stimulating interferon-γ (IFN-γ) production [46].
TDEVs mediate the activation of T cells and NK cells, driving a powerful anti-tumor effect. TDEVs loaded with the miRNAs Let-7i, miR-142, and miR-155 potentiate cytotoxic T cell activity, consequently inhibiting tumor progression in a murine model [47]. Moreover, TDEVs can enhance the antigen-presenting capacity of DCs, thereby improving anti-tumor efficacy. Specifically, EVs released from genetically engineered tumor cells exhibit high immunogenicity, which promotes DC maturation and further augments anti-tumor activity [48].
In summary, EVs contribute to immune activation in the TME by promoting M1 macrophage polarization, activating anti-tumor immune effectors, and facilitating antigen presentation, resulting in a potent anti-tumor outcome.
However, as tumors progress and the TME evolves, EVs contribute to the formation of an immunosuppressive microenvironment dominated by M2 [30]. For example, tissue inhibitor of metalloproteinase 3 (TIMP3) is a well-established tumor suppressor. In hepatocellular carcinoma (HCC), Hu et al. demonstrated that TDEVs facilitate M2 macrophage polarization by delivering miR-452-5p to downregulate TIMP3 expression, thereby accelerating malignant progression through the establishment of an immunosuppressive niche [49].
EVs from various cellular origins function as potent immunosuppressive agents, not only directly inhibiting T cell activity but also promoting their exhaustion, thereby dampening the overall anti-tumor immune response. M2 macrophage-derived EVs deliver miR-29a-3p and miR-21-5p to CD4+ T cells, activating signal transducer and activator of transcription 3 signaling and shifting the Treg/Th17 balance toward immunosuppression [50,51]. Ramil et al. found that immunosuppressive myeloid-derived suppressor cells (MDSCs) educated by CAFs release EVs containing fructose bisphosphatase 1, directly inhibiting T cell function [52]. The release of Treg-derived EVs (Treg-EVs) further reinforce the immunosuppressive microenvironment by polarizing macrophages toward an M2 phenotype, promoting the formation of tolerogenic DCs, and inhibiting the proliferation and activation of effector T cells [53].
The effects of TDEVs on immune cells are not a binary opposition of activation and suppression but are instead highly context-dependent. For instance, EVs engineered from myeloid leukemia cells to express the immunostimulatory ligands IL-15, IL-18, and 4-1BBL initially activate and enhance the proliferation of NK cells, thereby promoting anti-tumor activity. However, prolonged exposure for 48 h paradoxically suppresses the cytotoxic function of these NK cells [54].
The TME harbors EVs that display an immunomodulatory duality, capable of both activating anti-tumor immunity and suppressing immune cells. This duality offers a strategic avenue for developing next-generation immunotherapies aimed at amplifying the beneficial signals and blocking the inhibitory ones carried by EVs.
2.4 Crosstalk of EVs and Stromal Cells in the TME
CAFs represent the most abundant stromal population within the TME, playing an important role in angiogenesis, tumor growth and metastasis, and maintaining an immunosuppressive microenvironment by engaging in extensive intercellular crosstalk within the TME [55]. In cancer, CAFs can be transformed from normal fibroblasts [56], mesenchymal stem cells [57], adipocytes [58], and other cells.
EVs in the TME, particularly TDEVs, exhibit multifaceted effects on CAFs. Emerging studies have reported that TDEVs can induce the activation of lung fibroblasts to facilitate metastatic progression. Our team reported that Nidogen 1 in EVs derived from metastatic liver cancer cells is associated with the activation of lung fibroblasts, thereby enhancing tumor cell growth and invasion and pulmonary colonization [15]. Coincidentally, CAF-derived EVs have also demonstrated similar results. In salivary adenoid cystic carcinoma, Kong et al. found that CAF-derived EVs carrying ITGα2β1 facilitate EV uptake by lung fibroblasts and subsequent niche formation [59]. These findings collectively highlight integrin-mediated EV targeting as a promising therapeutic avenue for metastatic intervention.
Given the striking heterogeneity of CAF populations, TDEVs serve as key effectors that drive the functional diversification of fibroblasts within the TME. For instance, breast cancer-derived exosomes containing survivin upregulate superoxide dismutase 1 (SOD1) in resident fibroblasts, promoting their conversion into tumor-promoting myofibroblasts that secrete pro-tumorigenic factors to enhance cancer proliferation and metastatic dissemination [56]. Furthermore, EV-mediated crosstalk between tumor cells and fibroblasts facilitates the emergence of specialized CAF subtypes, including inflammatory CAFs (iCAFs) [55,60] and vascular CAFs (vCAFs) [61], which contribute to tumor progression through distinct mechanisms.
Similarly, CAF-derived EVs influence tumor cell behavior by enhancing malignant progression, metastatic potential, and therapy resistance through multifaceted mechanisms.
2.5 Anti-Tumor Functions of EVs in the TME
While EVs are widely found to promote tumor progression, a growing body of evidence from both in vitro and in vivo studies demonstrates that EVs can also exert potent anti-tumor responses by inhibiting proliferation and inducing apoptosis in cancer cells.
For instance, EC-EVs deliver miR-503 to tumor cells, markedly suppressing their proliferative, migratory, and invasive capacities [62]. Studies employing a simulated TME model have elucidated that the apoptotic effect in pancreatic cancer cells is mediated by miR-145 delivered via EVs originating from tumor-associated stromal cells [63]. EVs secreted by NK cells carry anti-tumor miRNAs. These EVs can trigger caspase-mediated apoptosis and suppress proliferation in tumor cells [64].
Nevertheless, research on the anti-tumor effects of EVs remains limited. Most studies are confined to in vitro models of the TME, which fail to fully recapitulate the complexity of in vivo conditions. Advancing this research direction is crucial for harnessing the therapeutic potential of EVs, underscoring the need for more intensive research efforts in this field (Fig. 2).
EVs facilitate tumor metastasis via three core mechanistic pathways. First, they directly transfer functional proteins, such as P-glycoprotein (P-gp) [65] and survivin [28], to recipient cells, conferring immediate phenotypic changes including chemotherapy resistance and enhanced cell survival. Second, EVs deliver regulatory molecules like non-coding RNAs (e.g., miR-155) [24] and cytokines (e.g., TNF-α) [46] to recipient cells, where they induce transcriptional reprogramming and modulate long-term cellular behaviors such as metastatic capacity and EMT. Third, EVs carry functional proteins that act on endothelial cells and fibroblasts, promoting angiogenesis, disrupting endothelial barrier integrity, and fostering the formation of a pre-metastatic microenvironment, all of which collectively facilitate tumor dissemination. These EV-mediated effects converge on key signaling pathways, notably PI3K/Akt [34] and NF-κB [22], forming an integrated regulatory network that drives tumor progression.

Figure 2: Extracellular vesicles (EVs) mediate intercellular communication within the tumor microenvironment (TME). EVs, released by diverse cell types within the TME, serve as crucial mediators of intercellular communication. This EV-driven crosstalk contributes to tumor progression by facilitating apoptosis resistance, angiogenesis, drug resistance, and immunosuppression. Conversely, EVs can also exert tumor-suppressive effects through the activation of antitumor immunity and the induction of tumor cell apoptosis. TAM: Tumor-Associated Macrophage; DC: Dendritic Cell; CAF: Cancer-Associated Fibroblast.
Beyond their role in mediating intercellular communication, EV biogenesis and functionality are profoundly influenced by the dynamic characteristics of TME. This section will systematically review how key TME features influence EV production, cargo sorting, and subsequent biological effects.
3.1 TME with Hypoxia Characteristics
Hypoxia constitutes a hallmark of solid tumors, driven by a profound imbalance between accelerated cell proliferation and inadequate vascular oxygen delivery, forcing tumor cells to undergo metabolic reprogramming. Hypoxia is involved in various processes related to cancer progression and remains a major driver of tumor resistance and recurrence [66].
In response to the hypoxic TME, cells within the TME secrete more EVs to meet intercellular communication demands, thereby influencing tumor progression. However, the underlying mechanisms remain incompletely understood. While the balance between multivesicular bodies (MVB) degradation and secretion is a key yet underexplored regulator of EV release, hypoxia is known to tip this balance toward secretion [67]. Specifically, it promotes the intracellular trafficking and plasma membrane fusion of MVBs, thereby elevating EV secretion. HOX transcript antisense RNA (HOTAIR) is a long non-coding RNA (lncRNA) frequently upregulated in various cancers. Research by Yang et al. revealed that HOTAIR upregulates the expression of RAB35, a key regulator of intracellular MVB transport, thereby enhancing MVB trafficking to the plasma membrane. Beyond this, HOTAIR overexpression also promotes the co-localization of vesicle-associated membrane protein 3 (VAMP3) with SNAP23 on MVB membranes, a step that facilitates subsequent MVB-plasma membrane fusion. Finally, phosphorylation of SNAP23 acts as the terminal trigger in this cascade, directly driving the secretion of EVs [68]. Another potential mechanism is that hypoxia-mediated disruption of intracellular MVB trafficking blocks the lysosomal fusion and degradation pathway. This redirection of MVB fate consequently promotes increased EV secretion. For example, in hypoxic head and neck squamous cell carcinoma (HNSCC), HIF-1α-mediated transcriptional repression reduces the expression of ATP6V1A, an essential V-ATPase subunit required for lysosomal homeostasis. The resulting lysosomal dysfunction disrupts MVB-lysosome fusion, thus promoting elevated EV secretion [69].
Hypoxia profoundly remodels the molecular cargo of EVs to exert pro-tumorigenic effects on recipient cells. EVs derived from mesenchymal stem cells under hypoxic conditions are enriched with miR-21-5p, which promotes tumor cell proliferation, metastatic dissemination, and M2 macrophage polarization [70]. EVs secreted by hypoxia-polarized M2-like TAMs deliver circ_0003137 to induce EMT in glioblastoma cells, thereby driving tumor progression and metastasis [71]. In addition, TDEVs from hypoxic TME carry pro-angiogenic miRNAs, including miR-135b [72] and miR-23a [73], which stabilize HIF-1α in endothelial cells through multiple mechanisms, ultimately promoting tumor angiogenesis. Additionally, hypoxia within the TME promotes the polarization of TAMs to an M2 phenotype. These M2-type TAMs release EVs enriched with cytokines and growth factors, which activates endothelial cells, promoting angiogenesis [74,75].
While the hypoxic TME enhances tumor cell uptake of EVs [76], how hypoxia alters extracellular EVs transport remains unexplored. Taken together, hypoxia governs a sophisticated web of EV-based crosstalk in the TME, which facilitates tumor adaptation and remodeling. Thus, interfering with hypoxic signaling or its downstream pathways offers a viable approach for developing novel anti-tumor therapies that target EV biology.
3.2 TME with Acidic Characteristics
Metabolic reprogramming drives tumor cells to secrete excessive lactate, whose accumulation establishes an acidic TME [77]. Recent studies have demonstrated that acidic TME orchestrates multifaceted pro-tumor effects, including suppression of immune cell function [78], evasion of immune surveillance [79], stimulation of angiogenesis [80], and induction of therapy resistance [81]. Collectively, these processes culminate in tumor invasion and metastasis.
Intercellular communication via EVs is profoundly sensitive to TME acidification, which alters vesicle biogenesis, transport, and uptake kinetics. First, an acidic microenvironment drives increased biogenesis, cargo reprogramming and secretion of EVs. In melanoma, Boussadia et al. demonstrated that in acidic TME, TDEVs are enriched with pro-invasive proteins, which confer heightened metastatic potential to tumor cells under neutral pH conditions [82]. Furthermore, an in vitro study indicates that acidic culture medium increases the yield of both proteins and nucleic acids in EVs. In contrast, alkaline conditions, or the use of proton pump inhibitors, suppress the secretion of EVs and reduce their molecular cargo [83]. However, the mechanism by which an acidic TME promotes EV biogenesis remains unclear. One hypothesis suggests that acidic TME accelerates the cellular turnover of lysosomes and EVs, thereby increasing the rates of exosome synthesis and release [84].
Acidic conditions within the TME promote EV uptake by modulating membrane composition and properties to facilitate efficient membrane fusion between EVs and recipient cells. From a biophysical perspective, the acidic conditions reduce charge shielding on the EV membrane, thereby decreasing electrostatic repulsion during fusion. Simultaneously, the stiffness of the EV membrane may become more comparable to that of the plasma membrane, further promoting fusion [85]. Moreover, changes in membrane lipid and protein composition are critical for efficient fusion between EVs and the cellular membrane. In a melanoma study, Parolini et al. demonstrated that an acidic environment not only enhances EV secretion and uptake but also alters EV membrane properties, resulting in high rigidity and elevated sphingomyelin/ganglioside GM3 content, features that may account for the improved fusion efficiency [86].
Targeting or reversing the tumor-promoting effects of the acidic TME represents a promising therapeutic strategy. The acidic TME fosters immune evasion by upregulating IFN-γ-dependent PD-L1 expression on tumor cells through STAT1/eIF4F translational activation. Neutralization of extracellular acidity has been shown to reduce PD-L1 expression while enhancing immune cell infiltration, thereby restoring antitumor immunity [87]. Building on the finding that acidic conditions promote EV uptake, a follow-up study reports the development of a smart drug delivery system specifically designed to leverage this phenomenon, which has substantially enhanced the effectiveness of tumor treatment [88]. Consequently, therapeutic interventions that either normalize the acidic microenvironment or target acid-regulated EV functions may offer a viable pathway to enhance treatment efficacy.
Therapeutic pressures, including chemotherapy and radiotherapy, drive tumor cells to dynamically remodel their EV secretion, thereby modulating intercellular communication within the stressed TME. This process reduces intracellular levels of toxic byproducts or drugs, while enabling the export of oncogenic molecules, thereby contributing to acquired drug resistance. For instance, both photodynamic therapy and chemotherapies trigger the rapid release of large quantities of EVs loaded with drugs, oncoproteins, and nucleic acids. This not only diminishes treatment efficacy but may also disseminate harmful cargo to distant healthy cells, exacerbating off-target effects [89]. Under cisplatin pressure, tumor cells package the drug into EVs for secretion, a direct efflux mechanism that lowers intracellular accumulation and promotes chemoresistance [89]. Similarly, irradiated cells secrete EVs carrying specific miRNAs that mediate bystander effects, facilitating communications between irradiated and non-irradiated cells [90]. Collectively, these findings highlight how therapy-stressed tumors exploit EV secretion to alter both local and systemic cell communication, supporting survival and compromising treatment outcomes.
In the TME, remodeling of the ECM and its mechanical properties influence cancer progression, a process that in turn regulates the biogenesis, secretion, and release of EVs from cancer cells [91]. For example, a stiffened ECM can enhance EV secretion by activating the Akt signaling pathway, which in turn upregulates Rab8 activity [92]. Nutrient deprivation imposes considerable stress on tumor cells, leading to increased EV secretion and altered cargo composition [93]. However, research in this area remains limited, and the specific mechanisms by which the ECM regulates EV biogenesis and extracellular transport are still not well understood. Further investigation into these processes is needed to clarify their role in tumor progression and therapy resistance.
Thus, the defining characteristics of the TME—hypoxia, acidity, therapeutic pressure, and ECM remodeling—converge to modulate EV biogenesis and function, profoundly influencing oncogenesis and tumor progression.
4 Therapeutic Potential and Frontier Prospect in Cancer
As discussed previously, EVs exhibit a dual role in the TME, capable of both promoting tumorigenesis and progression, and suppressing tumor advancement and metastasis. However, growing research attention has been focused on the intricate crosstalk mediated by EVs between tumor cells and immune cells. TDEVs can modulate the crosstalk among various immune cells within the TME to facilitate cancer growth and metastasis [94,95]. These mechanisms include impairing the differentiation and maturation of dendritic cells, suppressing the cytotoxicity of NK cells, and upregulating the population of Tregs, among others [96]. In-depth research into the mechanisms by which EVs participate in tumor immune responses will provide a more robust foundation for developing and applying effective strategies for cancer immunotherapy.
4.1 Potential of EVs in Cancer Treatment
The immunosuppressive tumor microenvironment represents a critical driver of malignant progression. To restore a normal immune environment, immunotherapy aims to restore and enhance immune system function, thereby killing tumor cells without harming the body. However, clinical trial results indicate that immunotherapy yields clinical responses in only a subset of patients, while the majority of patients still experience limited therapeutic benefit due to immune escape and tumor heterogeneity [97].
As mediators of intercellular crosstalk within the TME, EVs hold promise as tools for the development of immunotherapeutic strategies [98]. EVs’ inherent tumor targeting, biocompatibility, and barrier-crossing abilities have led to their initial application in immunotherapy. Given their ability to carry functional cargo, EV modification could allow them to deliver target molecules to recipient cells and exert their functions.
While immune checkpoint inhibitors (ICIs) have revolutionized cancer immunotherapy, their efficacy remains limited by the immunosuppressive TME [99]. Engineered EVs offer a promising alternative for immune checkpoint blockade, capable of restoring anti-tumor immunity with improved safety profiles. PD-1/PD-L1 axis is the star immune checkpoints in immunotherapy, as its interaction enables tumor cells to evade T cell-mediated immunity [100]. Although monoclonal antibodies against PD-1/PD-L1 axis can disrupt this interaction, they are associated with significant adverse effects and rapid clearance [101,102]. In recent years, many scientists have attempted to enable EVs expressing PD-1 from T cells, macrophages, and tumor cells, which can induce tumor cell apoptosis and block PD-L1-induced immune escape [103,104]. Furthermore, combination strategies employing EV-based ICIs with chemotherapy exhibit enhanced therapeutic effects [105]. This therapeutic strategy can also be applied to the interaction between CD47 on the surface of tumor cells and SIRPα on the surface of macrophages [106]. The inherent characteristics of engineered EVs make it safer and have fewer side effects in the field of blocking immune checkpoints, and they have broad prospects in the promotion and application of ICI therapy.
Tumor vaccines represent a promising immunotherapeutic strategy designed to activate robust immune responses and achieve anti-tumor efficacy through delivery of tumor-specific antigens and adjuvants [107]. Owing to their superior capacity for antigen presentation, DCs have been extensively utilized in the design of tumor vaccines to prime robust T-cell-mediated anti-tumor responses. EVs secreted by DCs carry MHC peptide complexes, co-stimulatory molecules, and adhesion molecules, and possess the same antigen-presenting capacity as DCs [108]. Compared to whole-cell vaccines, EV-based vaccines exhibit superior resistance to immunosuppressive signals in the TME, making them particularly effective for T cell activation. Lu et al. found that DC-derived EVs carrying alpha-fetoprotein activated CD8+ cytotoxic T lymphocytes in HCC mouse model, inducing a potent anti-tumor immune response and significantly inhibiting tumor progression [109]. Combining DC-derived EVs with ICIs further amplifies the therapeutic efficacy of DC-derived EVs, representing a promising combinatorial immunotherapeutic approach [110].
Related clinical trials are ongoing. In a metastatic melanoma trial, autologous DC-generated EVs loaded with tumor antigens demonstrated safety and feasibility, though no significant T cell responses were observed. Subsequent analysis suggested the observed anti-tumor activity may stem from NK cell activation and enhanced tumor infiltration [111]. However, the safety and efficacy of DC-derived EVs as tumor vaccines require further study. More recently, a melanoma trial utilizing plasmacytoid DC (pDC)-derived EV vaccines administered via subcutaneous injection showed both safety and the capacity to induce robust T cell-mediated anti-tumor immunity [112]. These contrasting outcomes highlight the need for further investigation into the safety profiles and therapeutic efficacy of DC-EV vaccines.
Beyond DCs, TDEVs themselves represent promising platforms for vaccine development. In pancreatic ductal adenocarcinoma (PDAC), engineered EVs from immunogenic dying tumor cells carrying CCL22 siRNA effectively disrupt the CCR4/CCL22 axis between DCs and Tregs, thereby inhibiting Treg expansion and preventing tumor growth. In combination with first-line chemotherapeutic drugs, these modified EVs demonstrate synergistic anti-tumor effects [113]. Huang et al. developed an engineered TDEVs rich in α-lactalbumin and introduced immunogenic cell death (ICD) inducers and TLR3 agonists. This engineered EV can induce cancer cell ICD, thereby releasing tumor antigens and TLR3 agonists. It promotes the activation of type one conventional DCs and CD8 T cell-mediated anti-tumor responses, thereby inhibiting the growth of triple-negative breast cancer tumors [114].
EVs have emerged as superior drug delivery vehicles due to their excellent biocompatibility, targeting properties, and cargo-protective capabilities [18]. Various therapeutic drugs can be loaded onto EVs in vitro, including small molecule drugs, nucleic acids, proteins, and nanoparticles [115,116]. EV-packaged small molecule chemotherapy drugs exhibit enhanced targeting efficiency, anti-tumor effects, and bioactivity [116].
As discussed above, functional RNAs exert important biological functions through intercellular crosstalk via EVs. Consequently, the therapeutic potential of loading therapeutic nucleic acid molecules, such as miRNAs, siRNAs, and CRISPR-Cas9, into EVs is increasingly being explored. MSC-derived EVs have emerged as effective miRNA delivery vehicles for miRNA-based therapies, demonstrating precise tissue targeting in various cancers, and significant anti-tumor effects across multiple cancer types [117,118]. While siRNA therapeutics offer precise gene silencing capability, their clinical translation has been limited by poor delivery efficiency, rapid degradation, and off-target effects. The membrane structure and targeting properties of EVs can facilitate the delivery of siRNAs to target organs [119]. The synergistic use of multiple anti-tumor molecules can significantly enhance therapeutic efficacy. Encapsulating both target gene-targeting siRNAs and anti-tumor drugs within EVs exhibits superior anti-tumor efficacy compared to monotherapy [120].
EVs functionalized with diverse metal nanoparticles represent a sophisticated, emerging platform for precision tumor therapy. Superparamagnetic iron oxide nanoparticles (SPIONs) enable magnetic field-guided tumor accumulation, significantly enhancing the delivery efficiency of EV-encapsulated chemotherapeutics [121]. Gold nanoparticles confer photothermal properties, converting absorbed light into localized hyperthermia for tumor ablation [122]. In combination with anti-miR-21, SPIONs and gold nanoparticles loaded simultaneously into EVs can achieve efficient photothermal therapy and anti-tumor effects [123].
EV-based therapeutic strategies hold significant potential, as evidenced by a growing body of clinical research. Although current trials have shown good safety profiles with infrequent serious adverse events, efficacy outcomes have been limited. Most studies do not reach their predefined efficacy endpoints, and the rate of patient benefit remains low (Table 1).
Additionally, the translation of EV-based therapies faces several key challenges (Fig. 3). First, limited production yield hinders the scalability of EV-based treatments. Secondly, the absence of standardized methodologies for EV isolation and purification poses a major obstacle. The lack of a consensus protocol results in substantial batch-to-batch and inter-laboratory heterogeneity, thereby giving rise to poor reproducibility, inconsistent therapeutic efficacy, and a lack of robust quality controls, all of which hinder the scalable manufacturing and clinical translation of EV-based therapeutics. Third, endogenous and exogenously administered EVs differ significantly in their biodistribution. Systemically delivered exogenous EVs are often rapidly cleared or accumulate in organs like the liver and spleen, whereas endogenous EVs show different targeting preferences. Consequently, this mismatch may prevent designed EV therapeutics from reaching their target tissues effectively. Fourth, the immunogenic potential of EVs raises important safety considerations for clinical translation. Specifically, tumor-derived or allogeneic EVs may induce unintended immunomodulatory effects or, in some contexts, promote tumor progression. Finally, the translational path is further complicated by the current lack of rigorous regulatory frameworks, which fail to provide clear quality standards and application guidelines for EV-based therapeutics.

Figure 3: Challenges in the Clinical Translation of Extracellular vesicle (EV)-Based Therapies. The clinical translation of EV-based therapies is hindered by several challenges. Key obstacles encompass limited production yields that impede scalable manufacturing, the absence of standardized methodologies undermining batch-to-batch reproducibility, and considerable heterogeneity in the biodistribution of EVs derived from different sources, which complicates predictable in vivo delivery. Furthermore, potential immunogenicity raises safety concerns regarding adverse reactions, while the lack of a clear regulatory system presents a significant barrier to clinical development and approval.
Table 1: Clinical trials involving EVs in cancer therapy.
| EVs Resources | Cargo | Disease | Outcomes | Phase | NCT Number/References | |
|---|---|---|---|---|---|---|
| Tumor vaccine | DCs | Tumor antigen | NSCLC | Not yet published | Phase II trial | NCT01159288 |
| Autologous DCs | MHC/peptide complexes | Metastatic melanoma | No grade II toxicity; MR: 1; PR:1; SD:1; PD: 1* | Phase I trial | [111] | |
| Plasmacytoid dendritic cells | MAGE 3 peptides | III/IV melanoma | Tolerated; no serious vaccine-induced side effects; PR:1 | Phase I trial | [112] | |
| DCs | IFN γ | NSCLC | Grade three hepatotoxicity; the median time to progression was 2.2 months and median overall survival was 15 months. | Phase II trial | [124] | |
| Autologous DCs | MAGE-A3, -A4, -A10, and MAGE-3DPO4 peptides | NSCLC | Tolerated with only grade 1–2 adverse events. | Phase I trial | [125] | |
| Drug delivery | Plant | Curcumin | Colon Cancer | Not yet published | Phase I trial | NCT01294072 |
| Mesenchymal stromal cells | KrasG12D siRNA | Metastatic pancreas cancer with KrasG12D mutation | Tolerated with no treatment-related adverse events and increase in intratumoral CD8+ T cells. | Phase I trial | NCT03608631 | |
| N.A. | STING agonist and PTGFRN | HNSCC, triple negative breast cancer, anaplastic thyroid carcinoma and cutaneous squamous cell carcinoma | Not yet published | Phase I/II trial | NCT04592484 | |
| N.A. | STAT6 anti-sense oligonucleotide | HCC and patients with liver metastases from either primary gastric cancer or colorectal cancer | Not yet published | Phase I trial | NCT05375604 |
Despite significant progress, most EV research is constrained by non-specific isolation methods that overlook the inherent heterogeneity of EVs by evaluating only their ensemble functions. Recent advances in single-vesicle analysis technologies have enabled detailed characterization of the heterogeneity in the size and surface phenotype of individual vesicles [126]. Furthermore, EVs exhibit significant heterogeneity in protein expression, suggesting that distinct EV subpopulations may perform markedly different functions [127]. However, current understanding of EVs function within TME is primarily derived from studying the collective behavior of total EVs isolated from specific cell lines. Omics sequencing and characterization at the single-vesicle level are therefore crucial for functional analysis and clinical translation of specific EV subsets [128]. Mirroring the rapid development of single-cell sequencing technologies, methods for EV classification and single-vesicle analysis, such as Droplet Digital Analysis, Nanoparticle Tracking Analysis (NTA), Interference Correlation Microscopy and high-sensitivity Flow Cytometry have also seen substantial progress [129,130]. These single-vesicle analysis platforms are now being explored for clinical diagnostics. The high-throughput analysis of EVs from serum or plasma samples has demonstrated significant potential for early disease detection [131,132]. Employing a high-sensitivity flow cytometry-based single-vesicle analysis platform, CD147-positive EV subpopulations can be specifically identified. This capability holds significant promise for advancing EV-based liquid biopsy strategies in both early detection and prognostic evaluation [133]. Infrared and Raman spectroscopy have been utilized to detect specific molecular signatures of prostate cancer-derived EVs, showing their potential utility in prostate cancer screening [134].
Although holding great promise, single-vesicle analysis technology is hampered by considerable technical challenges that hinder its clinical translation. The foremost challenge is cost, with current methodologies relying on costly instrumentation that impedes both clinical accessibility and broad adoption. Thus, developing highly sensitive, rapid, and economical detection platforms, including user-friendly microfluidic chips and portable biosensors, represents a critical unmet need for enabling point-of-care testing. Third, standardization remains a pressing issue that must be addressed for the advancement of single-vesicle analysis technology. Furthermore, the biological mechanisms underlying the heterogeneity of EVs revealed by single-vesicle analysis remain incompletely understood, warranting more focused research. Additionally, clinical studies validating its utility in early detection, diagnosis, and prognosis prediction are still lacking; more translational research is therefore needed to identify robust EV-based biomarkers and drive clinical implementation.
The advancement of single-vesicle analysis platforms is also poised to foster the advancement of EV-based cancer therapeutics. This technology advances the field by providing a more comprehensive profiling of tumor-associated EV signatures, thereby enhancing the accuracy of early detection and prognostic assessment. Simultaneously, it enables the detection of low-abundance EV features, which paves the way for developing personalized treatment strategies and targeted therapies.
EVs mediate complex intercellular crosstalk within TME through their diverse molecular cargo, including proteins, nucleic acids, and metabolites. These EV-transported biomolecules exhibit cargo-dependent functions, capable of both promoting and suppressing tumor progression. Furthermore, EVs carry tumor-specific information, making them promising diagnostic and translational biomarkers. A growing number of research is elucidating the biological mechanisms by which EVs promote tumor progression, providing a theoretical basis for the development of future therapeutic strategies.
EVs have emerged as transformative platforms for cancer therapy due to their biocompatibility, natural targeting capabilities, and remarkable stability in circulation. The combination of immune checkpoint blockers, tumor vaccines, anti-tumor drugs, and nucleic acid molecules with EVs has promising anti-tumor effects. Multifunctional, engineered EVs can even demonstrate superior therapeutic efficacy compared to single agents. Numerous clinical trials have been conducted. Nevertheless, the translation of EV-based therapeutics into clinical practice still confronts several challenges. These include the need for standardized manufacturing and quality control protocols, more efficient and cost-effective technologies, and further in-depth investigation. As these issues are addressed in the future, EV-based therapies are poised to demonstrate significant potential.
Acknowledgement:
Funding Statement: This study was supported by 0069/2023/ITP2, 0053/2024/RIB1, 0002/2025/NRP and 0008/2025/EQP from the Science and Technology Development Fund of the Macao and SRG2023-00042-ICMS from University of Macau.
Author Contributions: The authors confirm contribution to the paper as follows: Writing—original draft preparation and figures: Yaqi Xu; writing—review and editing: Jia Zhao and Xiaowen Mao; Conceptualization, supervision and finding acquisition: Xiaowen Mao. All authors reviewed and approved the final version of the manuscript.
Availability of Data and Materials: Not applicable.
Ethics Approval: Not applicable.
Conflicts of Interest: The authors declare no conflicts of interest.
References
1. Pitt JM , Kroemer G , Zitvogel L . Extracellular vesicles: masters of intercellular communication and potential clinical interventions. J Clin Investig. 2016; 126( 4): 1139– 43. doi:10.1172/JCI87316. [Google Scholar] [CrossRef]
2. Gu X , Chen A , Su Y , You M , Guo H , Tan S , et al. Extracellular vesicles: a new communication paradigm of complement in neurological diseases. Brain Res Bull. 2023; 199: 110667. doi:10.1016/j.brainresbull.2023.110667. [Google Scholar] [CrossRef]
3. Welsh JA , Goberdhan DCI , O’Driscoll L , Buzas EI , Blenkiron C , Bussolati B , et al. Minimal information for studies of extracellular vesicles (MISEV2023): from basic to advanced approaches. J Extracell Vesicles. 2024; 13( 2): e12404. doi:10.1002/jev2.12404. [Google Scholar] [CrossRef]
4. Tan Z , Yang C , Fu S , Wu J , Huang Y , Li H , et al. Migrasomes, critical players in intercellular communication. Cancer Cell Int. 2025; 25( 1): 113. doi:10.1186/s12935-025-03754-6. [Google Scholar] [CrossRef]
5. Jeppesen DK , Sanchez ZC , Kelley NM , Hayes JB , Ambroise J , Koory EN , et al. Blebbisomes are large, organelle-rich extracellular vesicles with cell-like properties. Nat Cell Biol. 2025; 27( 3): 438– 48. doi:10.1038/s41556-025-01621-0. [Google Scholar] [CrossRef]
6. Jeppesen DK , Fenix AM , Franklin JL , Higginbotham JN , Zhang Q , Zimmerman LJ , et al. Reassessment of exosome composition. Cell. 2019; 177( 2): 428– 45.e18. doi:10.1016/j.cell.2019.02.029. [Google Scholar] [CrossRef]
7. Mathieu M , Martin-Jaular L , Lavieu G , Théry C . Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat Cell Biol. 2019; 21( 1): 9– 17. doi:10.1038/s41556-018-0250-9. [Google Scholar] [CrossRef]
8. Yáñez-Mó M , Siljander PRM , Andreu Z , Bedina Zavec A , Borràs FE , Buzas EI , et al. Biological properties of extracellular vesicles and their physiological functions. J Extracell Vesicles. 2015; 4( 1): 27066. doi:10.3402/jev.v4.27066. [Google Scholar] [CrossRef]
9. Chan HY , Wang Q , Howie A , Bucci J , Graham P , Li Y . Extracellular vesicle biomarkers redefine prostate cancer radiotherapy. Cancer Lett. 2025; 616: 217568. doi:10.1016/j.canlet.2025.217568. [Google Scholar] [CrossRef]
10. Cargnoni A , Papait A , Masserdotti A , Pasotti A , Stefani FR , Silini AR , et al. Extracellular vesicles from perinatal cells for anti-inflammatory therapy. Front Bioeng Biotechnol. 2021; 9: 637737. doi:10.3389/fbioe.2021.637737. [Google Scholar] [CrossRef]
11. Wei Y , Wang D , Jin F , Bian Z , Li L , Liang H , et al. Pyruvate kinase type M2 promotes tumour cell exosome release via phosphorylating synaptosome-associated protein 23. Nat Commun. 2017; 8: 14041. doi:10.1038/ncomms14041. [Google Scholar] [CrossRef]
12. Bebelman MP , Janssen E , Pegtel DM , Crudden C . The forces driving cancer extracellular vesicle secretion. Neoplasia. 2021; 23( 1): 149– 57. doi:10.1016/j.neo.2020.11.011. [Google Scholar] [CrossRef]
13. Baglio SR , Lagerweij T , Pérez-Lanzón M , Ho XD , Léveillé N , Melo SA , et al. Blocking tumor-educated MSC paracrine activity halts osteosarcoma progression. Clin Cancer Res. 2017; 23( 14): 3721– 33. doi:10.1158/1078-0432.CCR-16-2726. [Google Scholar] [CrossRef]
14. Zhuang G , Wu X , Jiang Z , Kasman I , Yao J , Guan Y , et al. Tumour-secreted miR-9 promotes endothelial cell migration and angiogenesis by activating the JAK-STAT pathway. EMBO J. 2012; 31( 17): 3513– 23. doi:10.1038/emboj.2012.183. [Google Scholar] [CrossRef]
15. Mao X , Tey SK , Yeung CLS , Kwong EML , Fung YME , Chung CYS , et al. Nidogen 1-enriched extracellular vesicles facilitate extrahepatic metastasis of liver cancer by activating pulmonary fibroblasts to secrete tumor necrosis factor receptor 1. Adv Sci. 2020; 7( 21): 2002157. doi:10.1002/advs.202002157. [Google Scholar] [CrossRef]
16. Chalmin F , Ladoire S , Mignot G , Vincent J , Bruchard M , Remy-Martin JP , et al. Membrane-associated Hsp72 from tumor-derived exosomes mediates STAT3-dependent immunosuppressive function of mouse and human myeloid-derived suppressor cells. J Clin Investig. 2010; 120( 2): 457– 71. doi:10.1172/JCI40483. [Google Scholar] [CrossRef]
17. Wang J , Liu Q , Zhao Y , Fu J , Su J . Tumor cells transmit drug resistance via cisplatin-induced extracellular vesicles. Int J Mol Sci. 2023; 24( 15): 12347. doi:10.3390/ijms241512347. [Google Scholar] [CrossRef]
18. Marar C , Starich B , Wirtz D . Extracellular vesicles in immunomodulation and tumor progression. Nat Immunol. 2021; 22( 5): 560– 70. doi:10.1038/s41590-021-00899-0. [Google Scholar] [CrossRef]
19. Cavallari C , Camussi G , Brizzi MF . Extracellular vesicles in the tumour microenvironment: eclectic supervisors. Int J Mol Sci. 2020; 21( 18): 6768. doi:10.3390/ijms21186768. [Google Scholar] [CrossRef]
20. Tkach M , Théry C . Communication by extracellular vesicles: where we are and where we need to go. Cell. 2016; 164( 6): 1226– 32. doi:10.1016/j.cell.2016.01.043. [Google Scholar] [CrossRef]
21. Clancy JW , Sedgwick A , Rosse C , Muralidharan-Chari V , Raposo G , Method M , et al. Regulated delivery of molecular cargo to invasive tumour-derived microvesicles. Nat Commun. 2015; 6: 6919. doi:10.1038/ncomms7919. [Google Scholar] [CrossRef]
22. Zhang D , Zhang JW , Xu H , Chen X , Gao Y , Jiang HG , et al. Therapy-induced senescent tumor cell-derived extracellular vesicles promote colorectal cancer progression through SERPINE1-mediated NF-κB p65 nuclear translocation. Mol Cancer. 2024; 23( 1): 70. doi:10.1186/s12943-024-01985-1. [Google Scholar] [CrossRef]
23. Liu B , Shyr Y , Cai J , Liu Q . Interplay between miRNAs and host genes and their role in cancer. Brief Funct Genomics. 2019; 18( 4): 255– 66. doi:10.1093/bfgp/elz002. [Google Scholar] [CrossRef]
24. Li X , Chen Z , Ni Y , Bian C , Huang J , Chen L , et al. Tumor-associated macrophages secret exosomal miR-155 and miR-196a-5p to promote metastasis of non-small-cell lung cancer. Transl Lung Cancer Res. 2021; 10( 3): 1338– 54. doi:10.21037/tlcr-20-1255. [Google Scholar] [CrossRef]
25. Yang Q , Xu J , Gu J , Shi H , Zhang J , Zhang J , et al. Extracellular vesicles in cancer drug resistance: roles, mechanisms, and implications. Adv Sci. 2022; 9( 34): 2201609. doi:10.1002/advs.202201609. [Google Scholar] [CrossRef]
26. Ab Razak NS , Ab Mutalib NS , Mohtar MA , Abu N . Impact of chemotherapy on extracellular vesicles: understanding the chemo-EVs. Front Oncol. 2019; 9: 1113. doi:10.3389/fonc.2019.01113. [Google Scholar] [CrossRef]
27. Lv LH , Wan YL , Lin Y , Zhang W , Yang M , Li GL , et al. Anticancer drugs cause release of exosomes with heat shock proteins from human hepatocellular carcinoma cells that elicit effective natural killer cell antitumor responses in vitro. J Biol Chem. 2012; 287( 19): 15874– 85. doi:10.1074/jbc.M112.340588. [Google Scholar] [CrossRef]
28. Kreger BT , Johansen ER , Cerione RA , Antonyak MA . The enrichment of survivin in exosomes from breast cancer cells treated with paclitaxel promotes cell survival and chemoresistance. Cancers. 2016; 8( 12): 111. doi:10.3390/cancers8120111. [Google Scholar] [CrossRef]
29. Binenbaum Y , Fridman E , Yaari Z , Milman N , Schroeder A , Ben David G , et al. Transfer of miRNA in macrophage-derived exosomes induces drug resistance in pancreatic adenocarcinoma. Cancer Res. 2018; 78( 18): 5287– 99. doi:10.1158/0008-5472.can-18-0124. [Google Scholar] [CrossRef]
30. Dou X , Feng C , Li J , Jiang E , Shang Z . Extracellular vesicle-mediated crosstalk in tumor microenvironment dominates tumor fate. Trends Cell Biol. 2025; 35( 3): 230– 47. doi:10.1016/j.tcb.2024.08.008. [Google Scholar] [CrossRef]
31. Hüser L , Chhabra Y , Gololobova O , Wang V , Liu G , Dixit A , et al. Aged fibroblast-derived extracellular vesicles promote angiogenesis in melanoma. Cell Rep. 2024; 43( 9): 114721. doi:10.1016/j.celrep.2024.114721. [Google Scholar] [CrossRef]
32. Kalfon T , Loewenstein S , Gerstenhaber F , Leibou S , Geller H , Sher O , et al. Gastric cancer-derived extracellular vesicles (EVs) promote angiogenesis via angiopoietin-2. Cancers. 2022; 14( 12): 2953. doi:10.3390/cancers14122953. [Google Scholar] [CrossRef]
33. Yamada NO , Heishima K , Akao Y , Senda T . Extracellular vesicles containing microRNA-92a-3p facilitate partial endothelial-mesenchymal transition and angiogenesis in endothelial cells. Int J Mol Sci. 2019; 20( 18): 4406. doi:10.3390/ijms20184406. [Google Scholar] [CrossRef]
34. Han Y , Gong WS , Xing XS , Zhou H , Wang XL , Xu Y , et al. miR-183-5p-enriched extracellular vesicles promote the crosstalk between hepatocellular carcinoma cell and endothelial cell via SIK1/PI3K/AKT and CCL20/CCR6 signaling pathways. Front Oncol. 2025; 15: 1532239. doi:10.3389/fonc.2025.1532239. [Google Scholar] [CrossRef]
35. Lin J , Cao S , Wang Y , Hu Y , Liu H , Li J , et al. Long non-coding RNA UBE2CP3 enhances HCC cell secretion of VEGFA and promotes angiogenesis by activating ERK1/2/HIF-1α/VEGFA signalling in hepatocellular carcinoma. J Exp Clin Cancer Res. 2018; 37( 1): 113. doi:10.1186/s13046-018-0727-1. [Google Scholar] [CrossRef]
36. Xu ZH , Miao ZW , Jiang QZ , Gan DX , Wei XG , Xue XZ , et al. Brain microvascular endothelial cell exosome–mediated S100A16 up-regulation confers small-cell lung cancer cell survival in brain. FASEB J. 2019; 33( 2): 1742– 57. doi:10.1096/fj.201800428r. [Google Scholar] [CrossRef]
37. Nasir I , McGuinness C , Poh AR , Ernst M , Darcy PK , Britt KL . Tumor macrophage functional heterogeneity can inform the development of novel cancer therapies. Trends Immunol. 2023; 44( 12): 971– 85. doi:10.1016/j.it.2023.10.007. [Google Scholar] [CrossRef]
38. Wang L , Wang W , Hu D , Liang Y , Liu Z , Zhong T , et al. Tumor-derived extracellular vesicles regulate macrophage polarization: role and therapeutic perspectives. Front Immunol. 2024; 15: 1346587. doi:10.3389/fimmu.2024.1346587. [Google Scholar] [CrossRef]
39. Pavlov S , Ali E , Ambrozkiewicz F , Ye W , Rajtmajerová M , Liška V , et al. Macrophages in colorectal cancer: from normal mucosa to distant metastasis: beyond the M1/M2 paradigm. J Cancer. 2026; 17( 1): 157– 76. doi:10.7150/jca.126772. [Google Scholar] [CrossRef]
40. Ma RY , Black A , Qian BZ . Macrophage diversity in cancer revisited in the era of single-cell omics. Trends Immunol. 2022; 43( 7): 546– 63. doi:10.1016/j.it.2022.04.008. [Google Scholar] [CrossRef]
41. Wu SY , Fu T , Jiang YZ , Shao ZM . Natural killer cells in cancer biology and therapy. Mol Cancer. 2020; 19( 1): 120. doi:10.1186/s12943-020-01238-x. [Google Scholar] [CrossRef]
42. Eisenbarth SC . Dendritic cell subsets in T cell programming: location dictates function. Nat Rev Immunol. 2019; 19( 2): 89– 103. doi:10.1038/s41577-018-0088-1. [Google Scholar] [CrossRef]
43. Bao Q , Huang Q , Chen Y , Wang Q , Sang R , Wang L , et al. Tumor-derived extracellular vesicles regulate cancer progression in the tumor microenvironment. Front Mol Biosci. 2022; 8: 796385. doi:10.3389/fmolb.2021.796385. [Google Scholar] [CrossRef]
44. Moradi-Chaleshtori M , Bandehpour M , Heidari N , Mohammadi-Yeganeh S , Mahmoud Hashemi S . Exosome-mediated miR-33 transfer induces M1 polarization in mouse macrophages and exerts antitumor effect in 4T1 breast cancer cell line. Int Immunopharmacol. 2021; 90: 107198. doi:10.1016/j.intimp.2020.107198. [Google Scholar] [CrossRef]
45. Su MJ , Aldawsari H , Amiji M . Pancreatic cancer cell exosome-mediated macrophage reprogramming and the role of microRNAs 155 and 125b2 transfection using nanoparticle delivery systems. Sci Rep. 2016; 6: 30110. doi:10.1038/srep30110. [Google Scholar] [CrossRef]
46. Munich S , Sobo-Vujanovic A , Buchser WJ , Beer-Stolz D , Vujanovic NL . Dendritic cell exosomes directly kill tumor cells and activate natural killer cells via TNF superfamily ligands. Oncoimmunology. 2012; 1( 7): 1074– 83. doi:10.4161/onci.20897. [Google Scholar] [CrossRef]
47. Khani AT , Sharifzad F , Mardpour S , Hassan ZM , Ebrahimi M . Tumor extracellular vesicles loaded with exogenous Let-7i and miR-142 can modulate both immune response and tumor microenvironment to initiate a powerful anti-tumor response. Cancer Lett. 2021; 501: 200– 9. doi:10.1016/j.canlet.2020.11.014. [Google Scholar] [CrossRef]
48. Ito T , Sugiura K , Hasegawa A , Ouchi W , Yoshimoto T , Mizoguchi I , et al. Microbial antigen-presenting extracellular vesicles derived from genetically modified tumor cells promote antitumor activity of dendritic cells. Pharmaceutics. 2021; 13( 1): 57. doi:10.3390/pharmaceutics13010057. [Google Scholar] [CrossRef]
49. Hu Z , Chen J , Zhao Y , Yan C , Wang Y , Zhu J , et al. Exosomal miR-452-5p induce M2 macrophage polarization to accelerate hepatocellular carcinoma progression by targeting TIMP3. J Immunol Res. 2022; 2022: 1032106. doi:10.1155/2022/1032106. [Google Scholar] [CrossRef]
50. Clancy JW , D’Souza-Schorey C . Tumor-derived extracellular vesicles: multifunctional entities in the tumor microenvironment. Annu Rev Pathol. 2023; 18: 205– 29. doi:10.1146/annurev-pathmechdis-031521-022116. [Google Scholar] [CrossRef]
51. Zhou J , Li X , Wu X , Zhang T , Zhu Q , Wang X , et al. Exosomes released from tumor-associated macrophages transfer miRNAs that induce a treg/Th17 cell imbalance in epithelial ovarian cancer. Cancer Immunol Res. 2018; 6( 12): 1578– 92. doi:10.1158/2326-6066.CIR-17-0479. [Google Scholar] [CrossRef]
52. Ramil CP , Xiang H , Zhang P , Cronin A , Cabral L , Yin Z , et al. Extracellular vesicles released by cancer-associated fibroblast-induced myeloid-derived suppressor cells inhibit T-cell function. OncoImmunology. 2024; 13( 1): 2300882. doi:10.1080/2162402X.2023.2300882. [Google Scholar] [CrossRef]
53. Hou PP , Chen HZ . Extracellular vesicles in the tumor immune microenvironment. Cancer Lett. 2021; 516: 48– 56. doi:10.1016/j.canlet.2021.05.032. [Google Scholar] [CrossRef]
54. Li Q , Huang Q , Ting H , Wang Y , Huang Q , Shi J . Bifacial effects of engineering tumour cell-derived exosomes on human natural killer cells. Exp Cell Res. 2018; 363( 2): 141– 50. doi:10.1016/j.yexcr.2017.12.005. [Google Scholar] [CrossRef]
55. Kewitz-Hempel S , Windisch N , Hause G , Müller L , Sunderkötter C , Gerloff D . Extracellular vesicles derived from melanoma cells induce carcinoma-associated fibroblasts via miR-92b-3p mediated downregulation of PTEN. J Extracell Vesicles. 2024; 13( 9): e12509. doi:10.1002/jev2.12509. [Google Scholar] [CrossRef]
56. Li K , Liu T , Chen J , Ni H , Li W . Survivin in breast cancer–derived exosomes activates fibroblasts by up-regulating SOD1, whose feedback promotes cancer proliferation and metastasis. J Biol Chem. 2020; 295( 40): 13737– 52. doi:10.1074/jbc.RA120.013805. [Google Scholar] [CrossRef]
57. Purcell JW , Tanlimco SG , Hickson J , Fox M , Sho M , Durkin L , et al. LRRC15 is a novel mesenchymal protein and stromal target for antibody-drug conjugates. Cancer Res. 2018; 78( 14): 4059– 72. doi:10.1158/0008-5472.CAN-18-0327. [Google Scholar] [CrossRef]
58. Strong AL , Pei DT , Hurst CG , Gimble JM , Burow ME , Bunnell BA . Obesity enhances the conversion of adipose-derived stromal/stem cells into carcinoma-associated fibroblast leading to cancer cell proliferation and progression to an invasive phenotype. Stem Cells Int. 2017; 2017: 9216502. doi:10.1155/2017/9216502. [Google Scholar] [CrossRef]
59. Kong J , Tian H , Zhang F , Zhang Z , Li J , Liu X , et al. Extracellular vesicles of carcinoma-associated fibroblasts creates a pre-metastatic niche in the lung through activating fibroblasts. Mol Cancer. 2019; 18( 1): 175. doi:10.1186/s12943-019-1101-4. [Google Scholar] [CrossRef]
60. Wang D , Wang X , Song Y , Si M , Sun Y , Liu X , et al. Exosomal miR-146a-5p and miR-155-5p promote CXCL12/CXCR7-induced metastasis of colorectal cancer by crosstalk with cancer-associated fibroblasts. Cell Death Dis. 2022; 13( 4): 380. doi:10.1038/s41419-022-04825-6. [Google Scholar] [CrossRef]
61. Zhou Y , Ren H , Dai B , Li J , Shang L , Huang J , et al. Correction: hepatocellular carcinoma-derived exosomal miRNA-21 contributes to tumor progression by converting hepatocyte stellate cells tocancer-associated fibroblasts. J Exp Clin Cancer Res. 2022; 41( 1): 359. doi:10.1186/s13046-022-02575-z. [Google Scholar] [CrossRef]
62. Bovy N , Blomme B , Frères P , Dederen S , Nivelles O , Lion M , et al. Endothelial exosomes contribute to the antitumor response during breast cancer neoadjuvant chemotherapy via microRNA transfer. Oncotarget. 2015; 6( 12): 10253– 66. doi:10.18632/oncotarget.3520. [Google Scholar] [CrossRef]
63. Han S , Gonzalo DH , Feely M , Rinaldi C , Belsare S , Zhai H , et al. Stroma-derived extracellular vesicles deliver tumor-suppressive miRNAs to pancreatic cancer cells. Oncotarget. 2017; 9( 5): 5764– 77. doi:10.18632/oncotarget.23532. [Google Scholar] [CrossRef]
64. Cochran AM , Kornbluth J . Extracellular vesicles from the human natural killer cell line NK3.3 have broad and potent anti-tumor activity. Front Cell Dev Biol. 2021; 9: 698639. doi:10.3389/fcell.2021.698639. [Google Scholar] [CrossRef]
65. Lu JF , Luk F , Gong J , Jaiswal R , Grau GER , Bebawy M . Microparticles mediate MRP1 intercellular transfer and the re-templating of intrinsic resistance pathways. Pharmacol Res. 2013; 76: 77– 83. doi:10.1016/j.phrs.2013.07.009. [Google Scholar] [CrossRef]
66. Bai R , Li Y , Jian L , Yang Y , Zhao L , Wei M . The hypoxia-driven crosstalk between tumor and tumor-associated macrophages: mechanisms and clinical treatment strategies. Mol Cancer. 2022; 21( 1): 177. doi:10.1186/s12943-022-01645-2. [Google Scholar] [CrossRef]
67. He G , Peng X , Wei S , Yang S , Li X , Huang M , et al. Exosomes in the hypoxic TME: from release, uptake and biofunctions to clinical applications. Mol Cancer. 2022; 21( 1): 19. doi:10.1186/s12943-021-01440-5. [Google Scholar] [CrossRef]
68. Yang L , Peng X , Li Y , Zhang X , Ma Y , Wu C , et al. Long non-coding RNA HOTAIR promotes exosome secretion by regulating RAB35 and SNAP23 in hepatocellular carcinoma. Mol Cancer. 2019; 18( 1): 78. doi:10.1186/s12943-019-0990-6. [Google Scholar] [CrossRef]
69. Wang X , Wu R , Zhai P , Liu Z , Xia R , Zhang Z , et al. Hypoxia promotes EV secretion by impairing lysosomal homeostasis in HNSCC through negative regulation of ATP6V1A by HIF-1α. J Extracellular Vesicle. 2023; 12( 2): e12310. doi:10.1002/jev2.12310. [Google Scholar] [CrossRef]
70. Ren W , Hou J , Yang C , Wang H , Wu S , Wu Y , et al. Extracellular vesicles secreted by hypoxia pre-challenged mesenchymal stem cells promote non-small cell lung cancer cell growth and mobility as well as macrophage M2 polarization via miR-21-5p delivery. J Exp Clin Cancer Res. 2019; 38( 1): 62. doi:10.1186/s13046-019-1027-0. [Google Scholar] [CrossRef]
71. Liu L , Wang R , Alifu A , Xiao Y , Liu Y , Qian C , et al. Hypoxia-driven M2-polarized macrophages facilitate the epithelial-mesenchymal transition of glioblastoma via extracellular vesicles. Theranostics. 2024; 14( 16): 6392– 408. doi:10.7150/thno.95766. [Google Scholar] [CrossRef]
72. Umezu T , Tadokoro H , Azuma K , Yoshizawa S , Ohyashiki K , Ohyashiki JH . Exosomal miR-135b shed from hypoxic multiple myeloma cells enhances angiogenesis by targeting factor-inhibiting HIF-1. Blood. 2014; 124( 25): 3748– 57. doi:10.1182/blood-2014-05-576116. [Google Scholar] [CrossRef]
73. Hsu YL , Hung JY , Chang WA , Lin YS , Pan YC , Tsai PH , et al. Hypoxic lung cancer-secreted exosomal miR-23a increased angiogenesis and vascular permeability by targeting prolyl hydroxylase and tight junction protein ZO-1. Oncogene. 2017; 36( 34): 4929– 42. doi:10.1038/onc.2017.105. [Google Scholar] [CrossRef]
74. Ghalehbandi S , Yuzugulen J , Pranjol MZI , Pourgholami MH . The role of VEGF in cancer-induced angiogenesis and research progress of drugs targeting VEGF. Eur J Pharmacol. 2023; 949: 175586. doi:10.1016/j.ejphar.2023.175586. [Google Scholar] [CrossRef]
75. Ansardamavandi A , Tafazzoli-Shadpour M . Tumour endothelial cells in cancer: chemo-physical crosstalk and angiogenic signalling in the tumour microenvironment. Cell Signal. 2026; 141: 112391. doi:10.1016/j.cellsig.2026.112391. [Google Scholar] [CrossRef]
76. Cerezo-Magaña M , Christianson HC , van Kuppevelt TH , Forsberg-Nilsson K , Belting M . Hypoxic induction of exosome uptake through proteoglycan-dependent endocytosis fuels the lipid droplet phenotype in glioma. Mol Cancer Res. 2021; 19( 3): 528– 40. doi:10.1158/1541-7786.MCR-20-0560. [Google Scholar] [CrossRef]
77. Wang JX , Choi SYC , Niu X , Kang N , Xue H , Killam J , et al. Lactic acid and an acidic tumor microenvironment suppress anticancer immunity. Int J Mol Sci. 2020; 21( 21): 8363. doi:10.3390/ijms21218363. [Google Scholar] [CrossRef]
78. Song J , Ge Z , Yang X , Luo Q , Wang C , You H , et al. Hepatic stellate cells activated by acidic tumor microenvironment promote the metastasis of hepatocellular carcinoma via osteopontin. Cancer Lett. 2015; 356( 2): 713– 20. doi:10.1016/j.canlet.2014.10.021. [Google Scholar] [CrossRef]
79. Choi SYC , Collins CC , Gout PW , Wang Y . Cancer-generated lactic acid: a regulatory, immunosuppressive metabolite? J Pathol. 2013; 230( 4): 350– 5. doi:10.1002/path.4218. [Google Scholar] [CrossRef]
80. Wang Y , Liang Y , Li M , Lu J , Zhou S , Yu Y , et al. Single-cell multi-omics reveals that FABP1 + renal cell carcinoma drive tumor angiogenesis through the PLG-PLAT axis under fatty acid reprogramming. Mol Cancer. 2025; 24( 1): 179. doi:10.1186/s12943-025-02377-9. [Google Scholar] [CrossRef]
81. Faes S , Duval AP , Planche A , Uldry E , Santoro T , Pythoud C , et al. Acidic tumor microenvironment abrogates the efficacy of mTORC1 inhibitors. Mol Cancer. 2016; 15( 1): 78. doi:10.1186/s12943-016-0562-y. [Google Scholar] [CrossRef]
82. Boussadia Z , Lamberti J , Mattei F , Pizzi E , Puglisi R , Zanetti C , et al. Acidic microenvironment plays a key role in human melanoma progression through a sustained exosome mediated transfer of clinically relevant metastatic molecules. J Exp Clin Cancer Res. 2018; 37( 1): 245. doi:10.1186/s13046-018-0915-z. [Google Scholar] [CrossRef]
83. Ban JJ , Lee M , Im W , Kim M . Low pH increases the yield of exosome isolation. Biochem Biophys Res Commun. 2015; 461( 1): 76– 9. doi:10.1016/j.bbrc.2015.03.172. [Google Scholar] [CrossRef]
84. Damaghi M , Tafreshi NK , Lloyd MC , Sprung R , Estrella V , Wojtkowiak JW , et al. Chronic acidosis in the tumour microenvironment selects for overexpression of LAMP2 in the plasma membrane. Nat Commun. 2015; 6: 8752. doi:10.1038/ncomms9752. [Google Scholar] [CrossRef]
85. Zhai Y , Fang J , He F , Qin Z , Liu J , Li S . Mechanical strength and biomechanics of extracellular vesicles. Nano Today. 2026; 66: 102905. doi:10.1016/j.nantod.2025.102905. [Google Scholar] [CrossRef]
86. Parolini I , Federici C , Raggi C , Lugini L , Palleschi S , De Milito A , et al. Microenvironmental pH is a key factor for exosome traffic in tumor cells. J Biol Chem. 2009; 284( 49): 34211– 22. doi:10.1074/jbc.M109.041152. [Google Scholar] [CrossRef]
87. Knopf P , Stowbur D , Hoffmann SHL , Hermann N , Maurer A , Bucher V , et al. Acidosis-mediated increase in IFN-γ-induced PD-L1 expression on cancer cells as an immune escape mechanism in solid tumors. Mol Cancer. 2023; 22( 1): 207. doi:10.1186/s12943-023-01900-0. [Google Scholar] [CrossRef]
88. Gong C , Zhang X , Shi M , Li F , Wang S , Wang Y , et al. Tumor exosomes reprogrammed by low pH are efficient targeting vehicles for smart drug delivery and personalized therapy against their homologous tumor. Adv Sci. 2021; 8( 10): 2002787. doi:10.1002/advs.202002787. [Google Scholar] [CrossRef]
89. Aubertin K , Silva AKA , Luciani N , Espinosa A , Djemat A , Charue D , et al. Massive release of extracellular vesicles from cancer cells after photodynamic treatment or chemotherapy. Sci Rep. 2016; 6: 35376. doi:10.1038/srep35376. [Google Scholar] [CrossRef]
90. Xu S , Wang J , Ding N , Hu W , Zhang X , Wang B , et al. Exosome-mediated microRNA transfer plays a role in radiation-induced bystander effect. RNA Biol. 2015; 12( 12): 1355– 63. doi:10.1080/15476286.2015.1100795. [Google Scholar] [CrossRef]
91. Ansardamavandi A , Tafazzoli-Shadpour M . The functional cross talk between cancer cells and cancer associated fibroblasts from a cancer mechanics perspective. Biochim Biophys Acta BBA Mol Cell Res. 2021; 1868( 11): 119103. doi:10.1016/j.bbamcr.2021.119103. [Google Scholar] [CrossRef]
92. Wu B , Liu DA , Guan L , Myint PK , Chin L , Dang H , et al. Stiff matrix induces exosome secretion to promote tumour growth. Nat Cell Biol. 2023; 25( 3): 415– 24. doi:10.1038/s41556-023-01092-1. [Google Scholar] [CrossRef]
93. Li J , Lee Y , Johansson HJ , Mäger I , Vader P , Nordin JZ , et al. Serum-free culture alters the quantity and protein composition of neuroblastoma-derived extracellular vesicles. J Extracell Vesicles. 2015; 4: 26883. doi:10.3402/jev.v4.26883. [Google Scholar] [CrossRef]
94. Li J , Yu S , Rao M , Cheng B . Tumor-derived extracellular vesicles: key drivers of immunomodulation in breast cancer. Front Immunol. 2025; 16: 1548535. doi:10.3389/fimmu.2025.1548535. [Google Scholar] [CrossRef]
95. Jie H , Gluszko AC , Whiteside TL . Crosstalk of tumor-derived extracellular vesicles with immune recipient cells and cancer metastasis. Cancers. 2026; 18( 2): 196. doi:10.3390/cancers18020196. [Google Scholar] [CrossRef]
96. Liu X , To KKW , Zeng Q , Fu L . Effect of extracellular vesicles derived from tumor cells on immune evasion. Adv Sci. 2025; 12( 12): 2417357. doi:10.1002/advs.202417357. [Google Scholar] [CrossRef]
97. Gardner A , Ruffell B . Dendritic cells and cancer immunity. Trends Immunol. 2016; 37( 12): 855– 65. doi:10.1016/j.it.2016.09.006. [Google Scholar] [CrossRef]
98. Xia W , Tan Y , Liu Y , Xie N , Zhu H . Prospect of extracellular vesicles in tumor immunotherapy. Front Immunol. 2025; 16: 1525052. doi:10.3389/fimmu.2025.1525052. [Google Scholar] [CrossRef]
99. Chen Q , Zhang Q , He L , Yang M , Wang H , Lin M , et al. T-cell hitchhiking nanodrug activates the cGAS-STING signaling pathway for enhanced cancer immunotherapy. Acta Biomater. 2025; 208: 442– 55. doi:10.1016/j.actbio.2025.10.044. [Google Scholar] [CrossRef]
100. Gong J , Chehrazi-Raffle A , Reddi S , Salgia R . Development of PD-1 and PD-L1 inhibitors as a form of cancer immunotherapy: a comprehensive review of registration trials and future considerations. J Immunother Cancer. 2018; 6( 1): 8. doi:10.1186/s40425-018-0316-z. [Google Scholar] [CrossRef]
101. Davda J , Declerck P , Hu-Lieskovan S , Hickling TP , Jacobs IA , Chou J , et al. Immunogenicity of immunomodulatory, antibody-based, oncology therapeutics. J Immunother Cancer. 2019; 7: 105. doi:10.1186/s40425-019-0586-0. [Google Scholar] [CrossRef]
102. Deveuve Q , Lajoie L , Barrault B , Thibault G . The proteolytic cleavage of therapeutic monoclonal antibody hinge region: more than a matter of subclass. Front Immunol. 2020; 11: 168. doi:10.3389/fimmu.2020.00168. [Google Scholar] [CrossRef]
103. Xu F , Jiang D , Xu J , Dai H , Fan Q , Fei Z , et al. Engineering of dendritic cell bispecific extracellular vesicles for tumor-targeting immunotherapy. Cell Rep. 2023; 42( 10): 113138. doi:10.1016/j.celrep.2023.113138. [Google Scholar] [CrossRef]
104. Qiu Y , Yang Y , Yang R , Liu C , Hsu JM , Jiang Z , et al. Activated T cell-derived exosomal PD-1 attenuates PD-L1-induced immune dysfunction in triple-negative breast cancer. Oncogene. 2021; 40( 31): 4992– 5001. doi:10.1038/s41388-021-01896-1. [Google Scholar] [CrossRef]
105. Chen Y , Wang L , Zheng M , Zhu C , Wang G , Xia Y , et al. Engineered extracellular vesicles for concurrent Anti-PDL1 immunotherapy and chemotherapy. Bioact Mater. 2021; 9: 251– 65. doi:10.1016/j.bioactmat.2021.07.012. [Google Scholar] [CrossRef]
106. Tang L , Yin Y , Cao Y , Fu C , Liu H , Feng J , et al. Extracellular vesicles-derived hybrid nanoplatforms for amplified CD47 blockade-based cancer immunotherapy. Adv Mater. 2023; 35( 35): 2303835. doi:10.1002/adma.202303835. [Google Scholar] [CrossRef]
107. Peng M , Mo Y , Wang Y , Wu P , Zhang Y , Xiong F , et al. Neoantigen vaccine: an emerging tumor immunotherapy. Mol Cancer. 2019; 18( 1): 128. doi:10.1186/s12943-019-1055-6. [Google Scholar] [CrossRef]
108. Fernández-Delgado I , Calzada-Fraile D , Sánchez-Madrid F . Immune regulation by dendritic cell extracellular vesicles in cancer immunotherapy and vaccines. Cancers. 2020; 12( 12): 3558. doi:10.3390/cancers12123558. [Google Scholar] [CrossRef]
109. Lu Z , Zuo B , Jing R , Gao X , Rao Q , Liu Z , et al. Dendritic cell-derived exosomes elicit tumor regression in autochthonous hepatocellular carcinoma mouse models. J Hepatol. 2017; 67( 4): 739– 48. doi:10.1016/j.jhep.2017.05.019. [Google Scholar] [CrossRef]
110. Phung CD , Pham TT , Nguyen HT , Nguyen TT , Ou W , Jeong JH , et al. Anti-CTLA-4 antibody-functionalized dendritic cell-derived exosomes targeting tumor-draining lymph nodes for effective induction of antitumor T-cell responses. Acta Biomater. 2020; 115: 371– 82. doi:10.1016/j.actbio.2020.08.008. [Google Scholar] [CrossRef]
111. Escudier B , Dorval T , Chaput N , André F , Caby MP , Novault S , et al. Vaccination of metastatic melanoma patients with autologous dendritic cell (DC) derived-exosomes: results of thefirst phase I clinical trial. J Transl Med. 2005; 3( 1): 10. doi:10.1186/1479-5876-3-10. [Google Scholar] [CrossRef]
112. Charles J , Chaperot L , Hannani D , Bruder Costa J , Templier I , Trabelsi S , et al. An innovative plasmacytoid dendritic cell line-based cancer vaccine primes and expands antitumor T-cells in melanoma patients in a first-in-human trial. OncoImmunology. 2020; 9( 1): 1738812. doi:10.1080/2162402X.2020.1738812. [Google Scholar] [CrossRef]
113. Zhou W , Chen X , Zhou Y , Shi S , Liang C , Yu X , et al. Exosomes derived from immunogenically dying tumor cells as a versatile tool for vaccination against pancreatic cancer. Biomaterials. 2022; 280: 121306. doi:10.1016/j.biomaterials.2021.121306. [Google Scholar] [CrossRef]
114. Huang L , Rong Y , Tang X , Yi K , Qi P , Hou J , et al. Engineered exosomes as an in situ DC-primed vaccine to boost antitumor immunity in breast cancer. Mol Cancer. 2022; 21( 1): 45. doi:10.1186/s12943-022-01515-x. [Google Scholar] [CrossRef]
115. Salarpour S , Forootanfar H , Pournamdari M , Ahmadi-Zeidabadi M , Esmaeeli M , Pardakhty A . Paclitaxel incorporated exosomes derived from glioblastoma cells: comparative study of two loading techniques. DARU J Pharm Sci. 2019; 27( 2): 533– 9. doi:10.1007/s40199-019-00280-5. [Google Scholar] [CrossRef]
116. Meng W , He C , Hao Y , Wang L , Li L , Zhu G . Prospects and challenges of extracellular vesicle-based drug delivery system: considering cell source. Drug Deliv. 2020; 27( 1): 585– 98. doi:10.1080/10717544.2020.1748758. [Google Scholar] [CrossRef]
117. Wu P , Wang M , Jin C , Li L , Tang Y , Wang Z , et al. Highly efficient delivery of novel miR-13896 by human umbilical cord mesenchymal stem cell-derived small extracellular vesicles inhibits gastric cancer progression by targeting ATG2A-mediated autophagy. Biomater Res. 2024; 28: 119. doi:10.34133/bmr.0119. [Google Scholar] [CrossRef]
118. Zhou P , Du X , Jia W , Feng K , Zhang Y . Engineered extracellular vesicles for targeted reprogramming of cancer-associated fibroblasts to potentiate therapy of pancreatic cancer. Signal Transduct Target Ther. 2024; 9( 1): 151. doi:10.1038/s41392-024-01872-7. [Google Scholar] [CrossRef]
119. Zhan Q , Yi K , Cui X , Li X , Yang S , Wang Q , et al. Blood exosomes-based targeted delivery of cPLA2 siRNA and metformin to modulate glioblastoma energy metabolism for tailoring personalized therapy. Neuro Oncol. 2022; 24( 11): 1871– 83. doi:10.1093/neuonc/noac071. [Google Scholar] [CrossRef]
120. Gong L , Chen Z , Feng K , Luo L , Zhang J , Yuan J , et al. A versatile engineered extracellular vesicle platform simultaneously targeting and eliminating senescent stromal cells and tumor cells to promote tumor regression. J Nanobiotechnol. 2024; 22( 1): 105. doi:10.1186/s12951-024-02361-3. [Google Scholar] [CrossRef]
121. Zhang J , Ji C , Zhang H , Shi H , Mao F , Qian H , et al. Engineered neutrophil-derived exosome-like vesicles for targeted cancer therapy. Sci Adv. 2022; 8( 2): eabj8207. doi:10.1126/sciadv.abj8207. [Google Scholar] [CrossRef]
122. Kesharwani P , Ma R , Sang L , Fatima M , Sheikh A , Abourehab MAS , et al. Gold nanoparticles and gold nanorods in the landscape of cancer therapy. Mol Cancer. 2023; 22( 1): 98. doi:10.1186/s12943-023-01798-8. [Google Scholar] [CrossRef]
123. Bose RJ , Kumar SU , Zeng Y , Afjei R , Robinson E , Lau K , et al. Correction to “tumor cell-derived extracellular vesicle-coated nanocarriers: an efficient theranostic platform for the cancer-specific delivery of anti-miR-21 and imaging agents”. ACS Nano. 2023; 17( 19): 19470. doi:10.1021/acsnano.3c08363. [Google Scholar] [CrossRef]
124. Besse B , Charrier M , Lapierre V , Dansin E , Lantz O , Planchard D , et al. Dendritic cell-derived exosomes as maintenance immunotherapy after first line chemotherapy in NSCLC. Oncoimmunology. 2015; 5( 4): e1071008. doi:10.1080/2162402X.2015.1071008. [Google Scholar] [CrossRef]
125. Morse MA , Garst J , Osada T , Khan S , Hobeika A , Clay TM , et al. A phase I study of dexosome immunotherapy in patients with advanced non-small cell lung cancer. J Transl Med. 2005; 3( 1): 9. doi:10.1186/1479-5876-3-9. [Google Scholar] [CrossRef]
126. Dragovic RA , Gardiner C , Brooks AS , Tannetta DS , Ferguson DJP , Hole P , et al. Sizing and phenotyping of cellular vesicles using Nanoparticle Tracking Analysis. Nanomed Nanotechnol Biol Med. 2011; 7( 6): 780– 8. doi:10.1016/j.nano.2011.04.003. [Google Scholar] [CrossRef]
127. Mizenko RR , Brostoff T , Rojalin T , Koster HJ , Swindell HS , Leiserowitz GS , et al. Tetraspanins are unevenly distributed across single extracellular vesicles and bias sensitivity to multiplexed cancer biomarkers. J Nanobiotechnol. 2021; 19( 1): 250. doi:10.1186/s12951-021-00987-1. [Google Scholar] [CrossRef]
128. Banijamali M , Höjer P , Nagy A , Hååg P , Gomero EP , Stiller C , et al. Characterizing single extracellular vesicles by droplet barcode sequencing for protein analysis. J Extracell Vesicles. 2022; 11( 11): 12277. doi:10.1002/jev2.12277. [Google Scholar] [CrossRef]
129. Nogueras-Ortiz CJ , Eren E , Yao P , Calzada E , Dunn C , Volpert O , et al. Single-extracellular vesicle (EV) analyses validate the use of L1 Cell Adhesion Molecule (L1CAM) as a reliable biomarker of neuron-derived EVs. J Extracell Vesicles. 2024; 13( 6): e12459. doi:10.1002/jev2.12459. [Google Scholar] [CrossRef]
130. Yan S , Zhang W , Li X , Dutta S , Castle AR , Liu Y , et al. Single extracellular vesicle detection assay identifies membrane-associated α-synuclein as an early-stage biomarker in Parkinson’s disease. Cell Rep Med. 2025; 6( 3): 101999. doi:10.1016/j.xcrm.2025.101999. [Google Scholar] [CrossRef]
131. Chen M , Lin S , Zhou C , Cui D , Haick H , Tang N . From conventional to microfluidic: progress in extracellular vesicle separation and individual characterization. Adv Healthc Mater. 2023; 12( 8): 2202437. doi:10.1002/adhm.202202437. [Google Scholar] [CrossRef]
132. Zhu J , Wu F , Li C , Mao J , Wang Y , Zhou X , et al. Application of single extracellular vesicle analysis techniques. Int J Nanomed. 2023; 18: 5365– 76. doi:10.2147/ijn.s421342. [Google Scholar] [CrossRef]
133. Tian Y , Ma L , Gong M , Su G , Zhu S , Zhang W , et al. Protein profiling and sizing of extracellular vesicles from colorectal cancer patients via flow cytometry. ACS Nano. 2018; 12( 1): 671– 80. doi:10.1021/acsnano.7b07782. [Google Scholar] [CrossRef]
134. Krafft C , Wilhelm K , Eremin A , Nestel S , von Bubnoff N , Schultze-Seemann W , et al. A specific spectral signature of serum and plasma-derived extracellular vesicles for cancer screening. Nanomed Nanotechnol Biol Med. 2017; 13( 3): 835– 41. doi:10.1016/j.nano.2016.11.016. [Google Scholar] [CrossRef]
Cite This Article
 Copyright © 2026 The Author(s). Published by Tech Science Press.
Copyright © 2026 The Author(s). Published by Tech Science Press.This work is licensed under a Creative Commons Attribution 4.0 International License , which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
 Downloads
Downloads
 Citation Tools
Citation Tools