
 Submit a Paper
Submit a Paper Propose a Special lssue
Propose a Special lssue Open Access
Open Access
ARTICLE

Seed Priming with MgCl2, CaCl2, and ZnCl2 as a Biofortification Based-Approach Induces Changes in Anise Seedlings Emergence
Department of Field Crops, Faculty of Agriculture, Ankara University, Ankara, 06110, Türkiye
* Corresponding Author: Sibel Day. Email:
(This article belongs to the Special Issue: Role of Biostimulants in the Alleviation of Biotic and Abiotic Stress)
Phyton-International Journal of Experimental Botany 2023, 92(8), 2461-2471. https://doi.org/10.32604/phyton.2023.029920
Received 16 March 2023; Accepted 05 May 2023; Issue published 25 June 2023
 View Full Text
View Full Text Download PDF
Download PDFAbstract
Aromatic and medicinal plant species having small seeds have field emergence problems due to low nutrient supply. Therefore, Pimpinella anisum seeds were hydro and osmoprimed with 100 mM MgCl2, CaCl2, and ZnCl2, for 2, 4, and 8 h each to compare their growth attributes during germination and seedling establishment stages. Nontreated seeds were used as control. Both hydro and osmo primed seeds were dried for 48 h before, they were sown in plastic trays in growth room conditions to see the impact of treatments on seedling emergence and growth. The maximum root length (12.90 cm), fresh weight (256.30 mg plant−1), and mean emergence time (MET) were recorded from 8 h MgCl2 primed seeds. Similarly, the maximum (97.50, and 98.00%) emergence percentage was observed using 8 h MgCl2 primed seeds and nontreated seeds (control treatment). The evaluation of parameters like chlorophyll contents and electrical conductivity showed the 8 h MgCl2 priming as the optimum treatment. The evaluation of parameters like chlorophyll contents and electrical conductivity showed the 8 h MgCl2 priming as the optimum treatment. The result suggests MgCl2 priming worked synergistically and improved seedling growth attributes under greenhouse conditions. The chlorophyll content ranged 25.94–35.69 SPAD unit. The highest chlorophyll content was obtained from the seedlings obtained from 4 h CaCl2 treatment, which were statistically similar to the chlorophyll contents of the seedlings obtained after 8 h MgCl2 treatment and nontreated seeds (control treatment). All other treatments showed inhibition in the chlorophyll contents and growth attributes of the seedlings. In conclusion, MgCl2 osmopriming treatments were significantly promotive and better compared to hydro-priming and osmopriming treatments including control treatment in terms of anise seeds germination and emergence.Keywords
Anise-Pimpinella anisum L. (Umbelliferae, Apiaceae) is an important flowering, medicinal and aromatic plant native to the Eastern Mediterranean and South Western Asian regions [1,2] that grows to around 40–50 cm with white or yellow flowers [3]. It has been cultivated in Egypt for the last 4,000 years; from where it spread to other parts of Europe and the Middle East [4]. The species belonging to the genus Pimpinella are represented by 27 taxa in the flora of Türkiye [5] and have multiple uses in ethnomedicinal systems. It is used singly or in combination with other herbs and taken as a carminative stomachic, stimulant, expectorant, antispasmodic, and antiseptic [6]. It is also used as an aromatic flavoring agent in desserts and alcoholic beverages [7]. The plants grow and establish well on fertile, warm soils soon after spring [8].
Anise seeds have very small endosperm, and face difficulties during early stages of growth. Therefore seeds face multiple abiotic and biotic stresses; which may inhibit their germination, along with growth and development of seedlings under natural conditions [9,10]. Several strategies in soil fertilization are being employed by farmers to improve germination and emergence that vary depending on the farmers. Most of the farmers always tend to apply an easy and cheap way of crop production. Seed pre-sowing osmopriming treatments are widely used as an easy way to biofortify seeds with nutrient elements to stimulate their germination, emergence [11] and inhibit the effects of external stresses [12].
Hydroprimed and osmoprimed seeds of Oryza sativa L. [13,14], Brassica juncea L. [15], and Lens culinaris Medik. [16,17] showed improvement in germination and stand establishment. Although, anise is used as a multifunctional medicinal crop like Nigella sativa [18] and Ocimum basilicum [19] the studies on germination, seedling establishment and agronomic practices are rare. There is no study regarding the strategy of treating MgCl2 (Magnesium chloride), CaCl2 (Calcium chloride), and ZnCl2 (Zinc chloride) to improve anise germination and protect them from stresses during germination. This study evaluated the metabolic stimuli induced changes in anise seeds due to MgCl2, CaCl2, and ZnCl2 based osmopriming for obtaining uniform seed germination and stands in fields that are difficult to obtain under natural conditions [20]. These stored mineral nutrients like magnesium, calcium, and zinc have importance especially when seedlings are under conditions of limited nutrients. Enrichment of seeds via seed priming with these nutrients could promote germination and seedling growth.
Magnesium (Mg) is an important macronutrient element that is required to carry out several biochemical and physiological processes in plants [21] during photosynthesis by binding to chloroplasts, participating in the light harvest in PSI and PSII [22]. It is also involved in carbon fixation by chloroplasts [23]. Mg is a micronutrient element and an environment-friendly biostimulant. Mg deficiency reduces the rate of the biomass with a disruption of CO2 fixation and generation of reactive oxygen species (ROS) ending up in cell damage [22]. It also acts as an enzyme cofactor activity with ATP [24] based on hydrolysis and synthesis [25]. It is an important constituent of chlorophyll molecules and the powerhouse behind photosynthesis.
Calcium (Ca) is one of the main components of cell structure and it takes a role in cell elongation, division, controlling nutrient absorption, and helping water absorption [26]. Furthermore, due to its role in cell structure, and signaling roles in plants; Ca2+ ions are important in charge, osmotic balance, and in seed germination. When the seeds are imbibed with the impact of rehydration, gibberellins cause Ca2+ flux into the cytosol and the expression of calmodulin (Ca2+ binding proteins); which are involved in signal transduction [27,28]. Alpha-amylase enzyme one of the Ca2+ metalloenzymes required at the beginning of germination [29,30], is the consequence of Ca2+ related post-imbibition metabolism.
Zinc (Zn) is crucial during seed germination and the early growth stage of plants until the root system gains the ability to nutrient uptake from the soil [31]. Zinc is a co-factor of various enzymes that take a role in the detoxification of ROS. Beyond that stage, the seeds containing low zinc show a delay in germination and poor seedling vigor [31,32].
Chlorides (Cl) prevent the accumulation of free amino acids and protect plants against diseases with easy management of intercellular transport of water [33]. It is understood that both Mg and Cl ions could improve quality and yields in cereals and other crops.
Therefore, the present study aimed to evaluate the impact of water, MgCl2, CaCl2, and ZnCl2 priming on anise seeds germination and seedling growth under controlled conditions.
2.1 Seed Treatments (Priming) and Emergence Tests
The experimental material consisted of the anise seeds purchased from an anise farmer from Denizli Province in 2021.
Fifty seeds were used in each treatment with 4 replications. Nontreated seeds were used as control. Each of the aliquots of 50 seeds was immersed in 50 ml distilled water (hydropriming), 100 mM solutions of MgCl2 (EC = 8.3 mS cm−1), CaCl2 (EC = 9.4 mS cm−1), and ZnCl2 (EC = 8.9 mS cm−1) at 20°C for 2, 4, and 8 h under dark conditions. The osmoprimed seeds were rinsed and washed with distilled water to remove the traces of MgCl2, CaCl2 and ZnCl2 on the seed surfaces. Thereafter, the surfaces of these seeds were dried and left at room temperature for 2 days (22 ± 1°C) to decrease moisture contents [34]. The seeds in each replicate were weighed before and after priming treatment to calculate their water uptake.
Four replicates of 50 seeds (50 × 4 = 200 seeds) were sown at a depth of 2 cm in the plastic trays (30 cm × 21 cm × 9 cm) containing peat and placed in a growth chamber (Sanyo versatile Growth chamber, Japan) at 20 ± 1°C 45 μM photons m−2 s−1 light for 16 h. The peat used in the study had a pH of 6.5 and EC of 40 mS m−1, the porosity of around 69% (v w−1).
The number of emerged seedlings (unfolding cotyledons on the surface) was counted daily for up to 25 days, along with the emergence percentages of the respective seedlings. The plants were irrigated with 50 ml water 8 times during 26 days of the experiment. The mean emergence time (days) was calculated according to the formula given below ISTA [35]:
n = number of cotyledons on the turf surface at time t
t = days from planting
Σn = final number of the cotyledons on the turf surface
Chlorophyll contents were measured on the 25th day. Shoot length, root length, seedling fresh weight, and dry weight were measured for all seedlings from each replicate after the 26th day. Fresh weights of seedlings were measured soon after harvest to avoid weight loss [36]. The dry weight of the seedlings was measured after drying the samples in an oven at 70°C for 48 h [37].
2.2 Chlorophyll Content Measurement
Chlorophyll measurements were done with SPAD-502 Plus (Konica Minolta) using five leaves per seedling. Ten seedlings from each replicate were used for sampling [37].
2.3 Electrical Conductivity (EC) Test
The electrical conductivity (EC) of four replicates of 50 seeds each of soaked anise seeds with distilled water and different osmotic solutions for each treatment was measured using a WTW Cond 314i model conductivity meter [38]. The results were expressed in µS cm−1 to take account of variability in different treatments.
2.4 Experimental Design and Statistical Analysis
The experiment was arranged in a completely randomized design with four replicates. Water uptake and emergence percentage data were subjected to arcsine transformation before carrying out an analysis of variance with MSTAT-C statistical software (Michigan State University, version 2.10). The differences among the means were compared with Duncan’s Multiple Range Test (p < 0.01 or p < 0.05).
Seed water uptake varied among treatments (p = 0.0000). Seeds soaked in water for 8 h had the highest water uptake (96.00%) while the minimum water uptake (56.24%) was observed in 4 h CaCl2 soaking (Table 1).
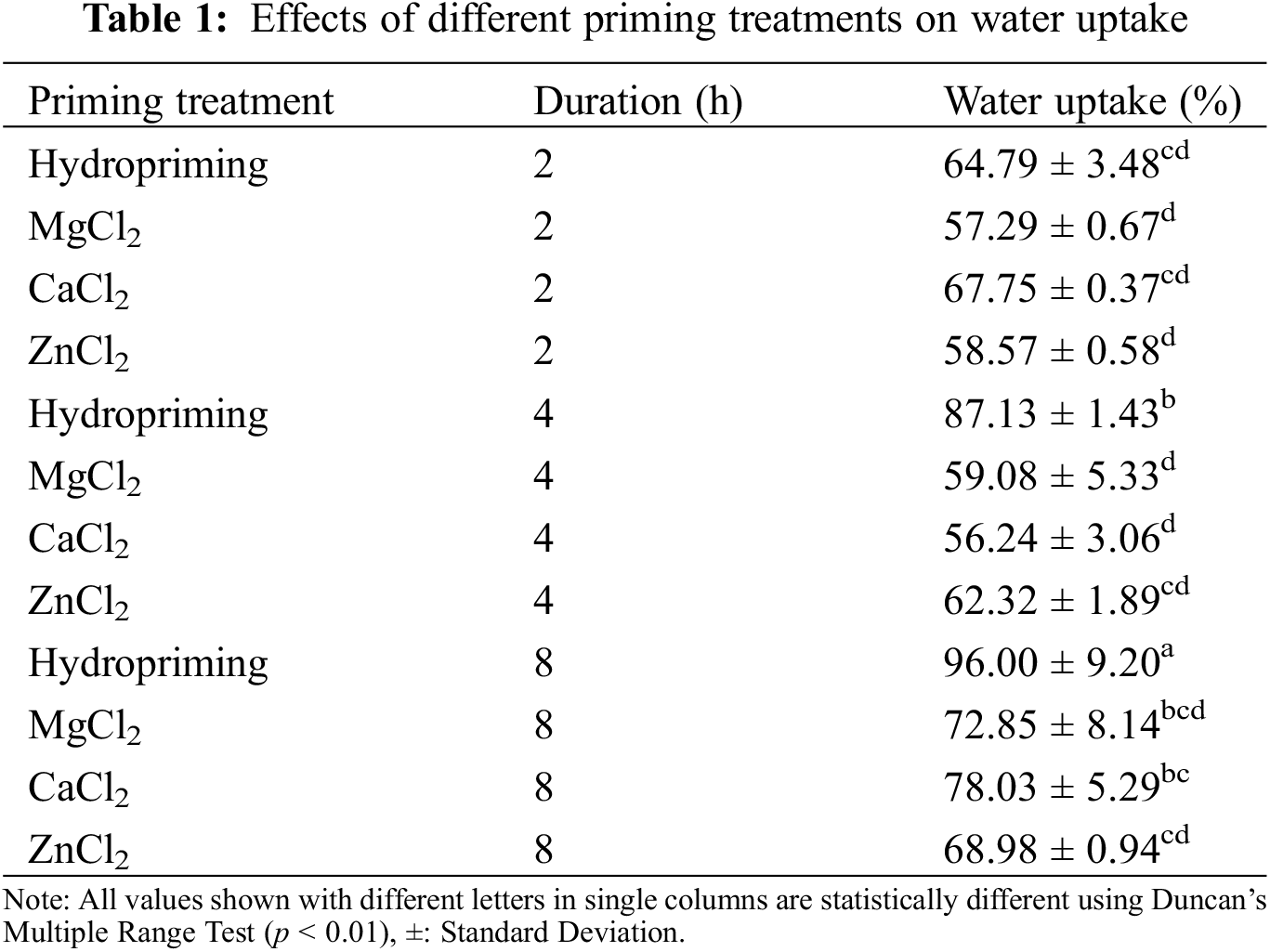
Mean emergence time represented a statistical difference over seed treatments (p < 0.0000). A comparison among seed priming treatments showed statistical differences among the non-treated, hydroprimed, and osmoprimed seeds. Non-primed seeds took 11.20 days to emerge. 8 h MgCl2 priming also shortened the mean emergence time over control, which was statistically different from non-treated control treatment and were statistically different among themselves showing a range of 9.73 to 17.09 days to emerge. The minimum and the maximum time to emergence were noted in 8 and 2 h MgCl2 priming. CaCl2 and ZnCl2 priming did not show any superiority over control or 8 h MgCl2 priming and the emergence time was slower (Table 2).
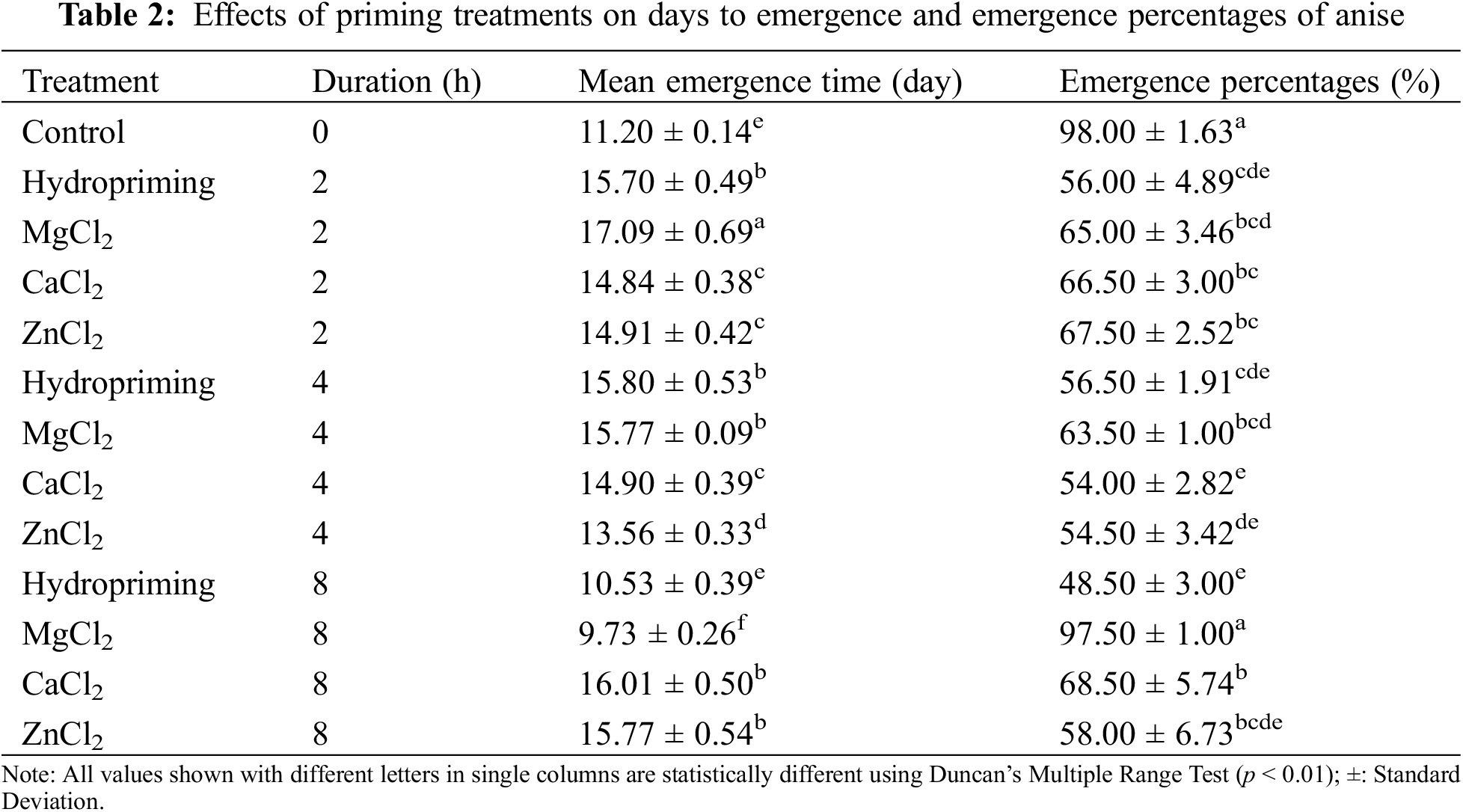
Impact of seed treatment on the emergence percentage showed statistically significant differences (p < 0.0000). The control and MgCl2 treatments showed maximum and statistically similar values of 98.00% and 97.50%, respectively. The hydroprimed seeds were negatively affected in terms of the emergence percentage compared to the non-treated seeds (control treatment). The minimum emergence percentage of 48.50% was noted on 8 h of hydroprimed seeds. Emergence percentages of CaCl2 primed seeds varied between 54.00% to 68.50% and ZnCl2 primed seeds varied between 54.50% to 67.50% (Table 2).
The root length of anise seedlings showed significant differences (p < 0.0000) among non-treated, hydroprimed, and osmoprimed seeds (Table 3). A comparison of the mean root length values showed that the maximum (12.90 cm) root length was noted from 8 h of MgCl2 primed seeds. Minimum root length was noted (5.43 cm) on 8 h ZnCl2 primed seeds. Hydroprimed and CaCl2 primed seeds had the longest roots in 8 and 2 h duration in the same order.
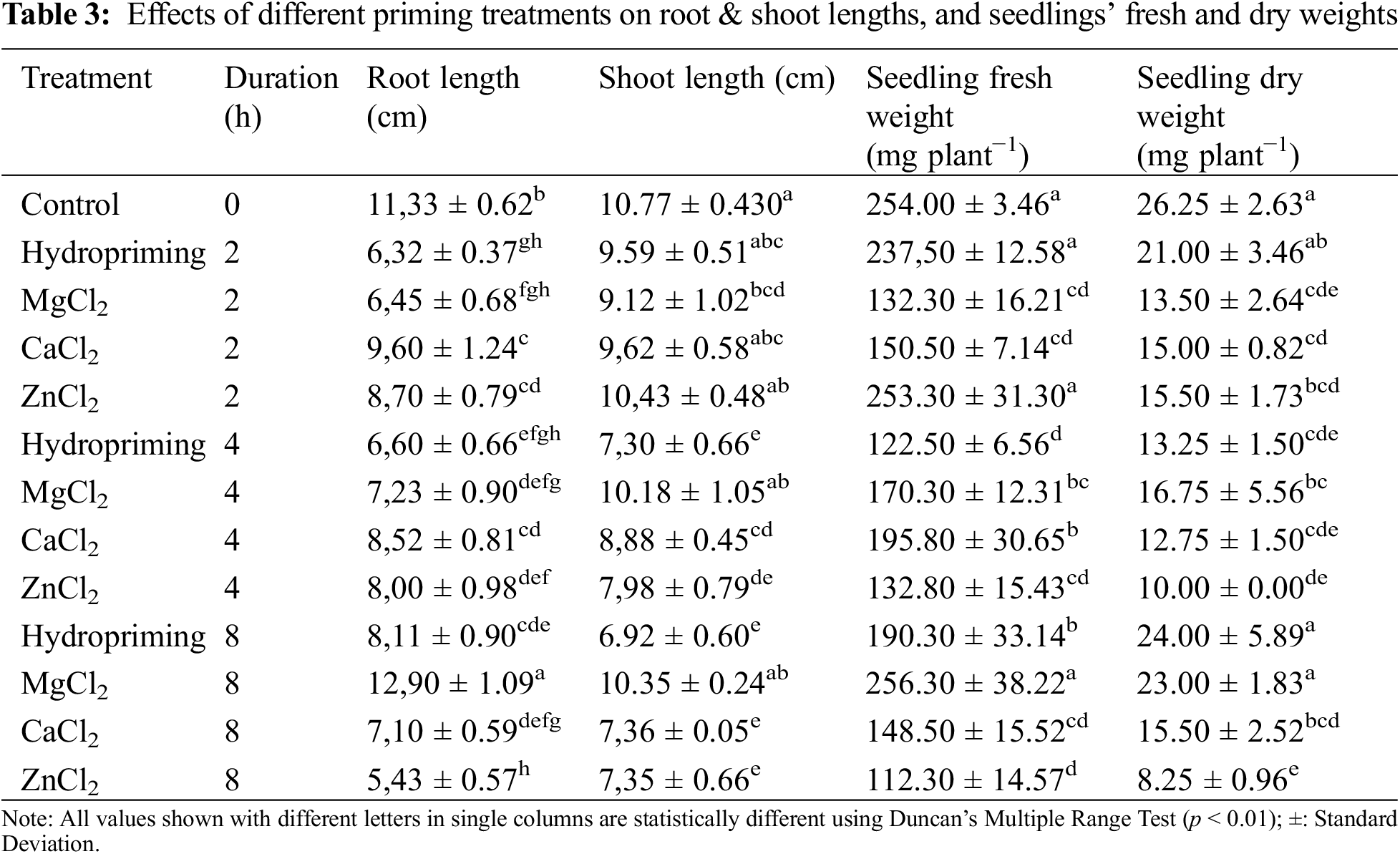
Analysis of variance showed significant differences (p < 0.0000) among treatments for shoot length. The maximum shoot length value of 10.77 cm was observed in the control (non-treated) treatment. This value was statistically similar to the 2 h hydroprimed seeds with values of 9.59 cm, 4 and 8 h MgCl2 primed seeds with values of 10.18, 10.35 cm and 2 h CaCl2 and ZnCl2 primed seeds with values of 9.62 and 10.43 cm. The minimum shoot length value of 6.92 cm was noted from 8 h hydroprimed seeds. It seemed that the negative effects of hydropriming on growth parameters were due to over-imbibition by anise seeds during hydropriming (Table 3).
Seedling fresh weight was significantly (p < 0.0000) affected by seed treatments (Table 3). The maximum seedling fresh weight value of 256.30 mg plant−1 was noted in 8 h MgCl2 treated seeds, which showed statistical similarity with non-treated seeds (254.00 mg plant−1), 2 h hydroprimed seeds (237.50 mg plant−1), and 2 h ZnCl2 treatment (253.30 mg plant−1). The lowest seedling fresh weights were obtained from 8 h ZnCl2 primed seeds with a value of 112.30 mg plant−1.
The seedling dry weight values of anise were significantly affected (p < 0.0000) by seed treatments. While the maximum seedling dry weight value was observed in non-treated seeds (26.25 mg plant−1), statistical similarity with 8 h hydropriming (24.00 mg plant−1) and 8 h MgCl2 (23.00 mg plant−1) treatment were also observed (Table 3).
Results about chlorophyll contents measurements (Table 4) clearly reflected the sharp impact (p < 0.0000) of seed treatments on chlorophyll contents. The maximum chlorophyll contents in terms of SPAD unit were observed in 4 h CaCl2 priming with 35.69 SPAD units and statistically similar values were noted in control (34.51 SPAD units) and 8 h MgCl2 priming (35.18 SPAD units). The osmoprimed seeds improved the chlorophyll contents depending on the duration of treatment.
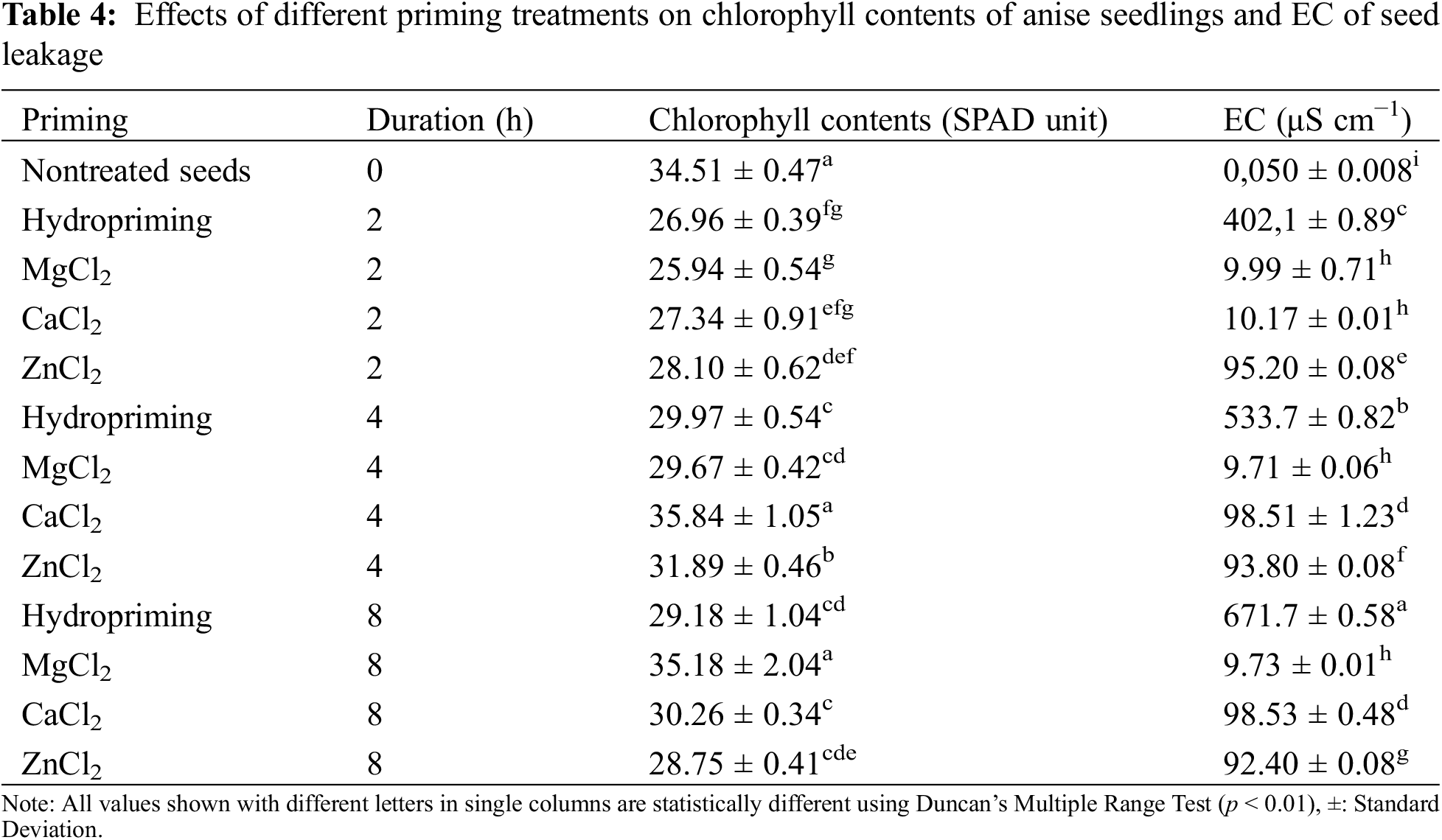
The electrical conductivity test showed significantly dissimilar variations (p < 0.0000). The minimum values were observed in MgCl2 priming in all treatment durations between 9.71–9.99 μS cm−1 (Table 4). The maximum EC value was observed in 8 h hydropriming with 671.7 μS cm−1.
During priming treatments, the seeds were partially hydrated with water and different osmotic agents to a point (2, 4, and 8 h). Water uptake showed diversity among treatments. The results revealed that among priming treatments hydropriming had the highest water uptake in all durations. This could be attributed to a low EC value of distilled water compared to other treatments, leading to uncontrolled water uptake.
MgCl2 priming treatments showed an increase in root length, fresh and dry weight along with chlorophyll contents with each increase in priming treatment duration of 2 to 8 h. CaCl2 priming treatment showed maximum improvement after 4 h of priming treatment. These two treatments were at par with the values obtained from nonprimed seeds used as control treatment. ZnCl2 priming treatments were noneffective at any duration of treatment and were inferior compared to the control treatment. The maximum values for these parameters were noted on 2 h treatment duration followed by obtaining decreased values for each parameter on 4 and 8 h priming duration treatments. The results indicated variable inhibition in these parameters for the rest of the primings for 2, 4, and 8 h excluding 8 h MgCl2 treatment.
Seedling dry weight in control had the maximum value among all treatments but no statistical difference was observed with 8 h MgCl2 priming. Priming duration in each treatment showed dissimilar results. This showed that the seeds could end up with either of the
a) replacement of seed nutrient matters in the cytoplasm with water moving from outside.
b) accumulation of the undesired amount of water into the seed cells.
c) bursting of cells, which are not desired beyond maintenance of osmotic balance between the internal and external forces.
An imbalance in the solid loss/water gain ratio is not desired and ends up in embryo damage with negative implications on the growth of seedlings’ roots and shoots depending on the environmental factors and genetic potential of the seeds [39,40].
Mg has an important role in the development and formation of sink organs [41,42] and the extra Mg given through osmopriming helped seed and seedling attributes to improve them in terms of fresh seedling weight per plant over other treatments. The nutrient matters are lost and washed away through osmosis with each increase in the duration of hydropriming, which influence the growth and development of roots from the seeds [42]. MgCl2 osmopriming maintains the genetic fidelity of the seeds, which should be carried out very carefully. Taking water up to a critical point is essential for germination and improving seed vigor, with better root growth and uniform crop establishment, and larger canopy [43,27]. This helps in improved competition with the surrounding plants and higher yields [44].
Zinc is important during germination against abiotic stress and for seedling vigor. However, ZnCl2 priming induced inhibition, toxicity, and dormancy during germination and growth compared to the control or other priming treatments. It seemed the amount and duration of ZnCl2 (100 mM) treatments used in the current experiment were toxic for anise. These findings were corroborated by Stanković et al. [45] who had similar findings using different levels of ZnCl2 to germinate wheat seeds germination observing limited seedling growth.
Hydropriming during 2, 4 and 8 h led to uncontrolled water uptake of anise seeds and less emergence percentage than other treatments. Imbibition damage was also evident due to minimum germination in 8 h hydropriming. Imbibition damage due to uncontrolled rapid water uptake causes cell death and high solute leakage which was observed as the highest in hydropriming compared to other treatments. Considering these results 8 h of priming with water reduced the emergence percentage for anise seeds. The disadvantages of hydropriming were mentioned by Lutts et al. [46] and it is also recommended to define accurate treatment duration, temperature, and water volume for every species. On the contrary osmopriming with 8 h MgCl2 created a positive impact on emergence percentage due to slow water uptake related to low water potential. Positive effects of MgCl2 seed priming on germination percentage have been noted in rice by Brooks et al. [47].
A decrease in chlorophyll content leads to low photosynthesis which has a negative impact on plant growth [37]. Magnesium is necessary for the synthesis of chlorophyll, and essential for photosynthesis [14] and its deficiency causes a reduction in chlorophyll concentrations in leaves [42]. Additionally, Wang et al. [48] have already mentioned the protective role of Ca2+ on photosynthetic electron transport. The results show similarity with the observation showing 8 h MgCl2 and 4 h CaCl2 priming of anise seeds led to an increase in chlorophyll synthesis compared to hydroprimed seeds in this research.
Electrolyte leakage has been reported as the determiner of seed viability and vigor in several species. The seeds having less vigor leak more electrolytes during imbibition [49,50]. The hydroprimed anise seeds had less emergence percentage and had the maximum EC values which seemed as an indicator of hydropriming with a negative impact on anise seeds and germination. However, priming with MgCl2 gave the minimum EC values among priming treatments, which was the evidence of repair mechanism of the membrane, especially for 8 h MgCl2 priming.
Sustaining favorable water under field conditions is critical for the germination process. Priming with different osmotic agents and water generated moderate abiotic stress (osmotic stress, saline, and drought stress) during soaking, and accumulation of osmotically active solutes like proline, which is reported in a dissimilar number of species during seed priming [51–53,46,42].
Any duration of hydropriming treatments is not recommended for anise. It seemed all treatment durations in hydropriming had inhibitory effects due to imbibition-based damages. This trend was also indicative in all osmoprimed seeds, excluding 8 h MgCl2 priming treated seeds. Although 8 h MgCl2 priming treatment and non-treated control treatment were statistically similar the former showed a numerical improvement over the other. This showed that biofortification of MgCl2 improved anise seed germination and emergence.
Funding Statement: The authors received no specific funding for this study.
Author Contributions: The authors confirm contribution to the paper as follows: study conception and design: Sibel Day, Nilüfer Koçak-Şahin; data collection: Sibel Day, Nilüfer Koçak-Şahin; analysis and interpretation of results: Sibel Day; draft manuscript preparation: Sibel Day. All authors reviewed the results and approved the final version of the manuscript.
Conflicts of Interest: The authors declare that they have no conflicts of interest to report regarding the present study.
References
1. Shojaii, A., Abdollahi Fard, M. (2012). Review of pharmacological properties and chemical constituents of Pimpinella anisum. International Scholarly Research Notices, 2012(3), 1–8. https://doi.org/10.5402/2012/510795 [Google Scholar] [PubMed] [CrossRef]
2. Thiviya, P., Gamage, A., Piumali, D., Merah, O., Madhujith, T. (2021). Apiaceae as an important source of antioxidants and their applications. Cosmetics, 8(4), 111. https://doi.org/10.3390/cosmetics8040111 [Google Scholar] [CrossRef]
3. Kara, N. (2015). Yield, quality, and growing degree days of anise (Pimpinella anisum L.) under different agronomic practices. Turkish Journal of Agriculture and Forestry, 39(6), 1014–1022. https://doi.org/10.3906/tar-1411-143 [Google Scholar] [CrossRef]
4. Boztas, G., Bayram, E. (2021). Determination of the agronomic, morphological and physiological differences which effected yield and quality in the developed anise lines. Journal of Agricultural Faculty of Bursa Uludag University, 35(1), 55–74 (in Turkish, with abstract in English). [Google Scholar]
5. TUBIVES (2023). Turkish plants data service. http://194.27.225.161/yasin/tubives/index.php [Google Scholar]
6. Shahrajabian, M. H., Sun, W., Cheng, Q. (2019). Chinese star anise and anise, magic herbs in traditional Chinese medicine and modern pharmaceutical science. Asian Journal of Medical and Biological Research, 5(3), 162–179. https://doi.org/10.3329/ajmbr.v5i3.43584 [Google Scholar] [CrossRef]
7. Sun, W., Shahrajabian, M. H., Cheng, Q. (2019). Anise (Pimpinella anisum L.a dominant spice and traditional medicinal herb for both food and medicinal purposes. Cogent Biology, 5(1), 1–25. https://doi.org/10.1080/23312025.2019.1673688 [Google Scholar] [CrossRef]
8. Katsoulis, G. I., Kimbaris, A. C., Anastasaki, E., Damalas, C. A., Kyriazopoulos, A. P. (2022). Chamomile and anise cultivation in olive agroforestry systems. Forests, 13(1), 128. https://doi.org/10.3390/f13010128 [Google Scholar] [CrossRef]
9. Kaya, M. D., Ozcan, F., Day, S., Bayramin, S., Ipek, A. (2013). Allelopathic role of essential oils in sunflower stubble on germination and seedling growth of the subsequent crop. International Journal of Agriculture and Biology, 15(2), 337–341. [Google Scholar]
10. Hoseini, A., Salehi, A., Sayyed, R. Z., Balouchi, H., Moradi, A. et al. (2022). Efficacy of biological agents and fillers seed coating in improving drought stress in anise. Frontiers in Plant Science, 13, 1–12. https://doi.org/10.3389/fpls.2022.955512 [Google Scholar] [PubMed] [CrossRef]
11. Hossain, M. A., Li, Z. G., Hoque, T. S., Burritt, D. J., Fujita, M. et al. (2018). Heat or cold priming-induced cross tolerance to abiotic stresses in plants: Key regulators and possible mechanism. Protoplasma, 255(1), 399–412. https://doi.org/10.1007/s00709-017-1150-8 [Google Scholar] [PubMed] [CrossRef]
12. Savvides, A., Ali, S., Tester, M., Fotopoulos, V. (2016). Chemical priming of plants against multiple abiotic stresses: Mission possible? Trends in Plant Science, 21(4), 329–340. https://doi.org/10.1016/j.tplants.2015.11.003 [Google Scholar] [PubMed] [CrossRef]
13. Prom-u-thai, C., Rerkasem, B., Yazıcı, A., Cakmak, I. (2012). Zinc priming promotes seed germination and seedling vigor of rice. Journal of Plant Nutrition and Soil Science, 175(3), 482–488. https://doi.org/10.1002/jpln.201100332 [Google Scholar] [CrossRef]
14. Kareem, I., Ismail, M. R., Pueth, A. (2019). Suitable priming for rice yield improvement. Cercetări Agronomice în Moldova, 52(1), 1–16. https://doi.org/10.2478/cerce-2019-0001 [Google Scholar] [CrossRef]
15. Srivastava, A. K., Lokhande, V. H., Patade, V. Y., Suprasanna, P., Sjahril, R. et al. (2010). Comparative evaluation of hydro-, chemo-, and hormonal priming methods for imparting salt and PEG stress tolerance in Indian mustard (Brassica juncea L.). Acta Physiolgiae Plantarum, 32(6), 1135–1144. https://doi.org/10.1007/s11738-010-0505-y [Google Scholar] [CrossRef]
16. Saglam, S., Day, S., Kaya, G., Gürbüz, A. (2010). Hydropriming increases germination of lentil (Lens culinaris Medik.) under water stress. Notulae Scientia Biologicae, 2(2), 103–106. https://doi.org/10.15835/nsb224602 [Google Scholar] [CrossRef]
17. Farooq, M., Romdhane, L., Al Sulti, M. K. R. A., Rehman, A., Al-Busaidi, W. M. et al. (2020). Morphological, physiological and biochemical aspects of osmopriming induced drought tolerance in lentil. Journal of Agronomy and Crop Science, 206(2), 176–186. https://doi.org/10.1111/jac.12384 [Google Scholar] [CrossRef]
18. Day, S., Abay, G., Özgen, Y., Önol, B. (2022). Effect of sulphur treatments on growth parameters and oil yield of black cumin (Nigella sativa L.). In: Gesunde pflanzen. https://doi.org/10.1007/s10343-022-00793-1 [Google Scholar] [CrossRef]
19. Day, S. (2021). Secondary metabolites of Ocimum basilicum L. In: Walton, A. A. (Ed.Ocimum basilicum: Taxonomy, cultivation and uses, pp. 1–30. Newyork: Nova Science. [Google Scholar]
20. Kumar, H. D., Aloke, P. (2020). Role of biostimulant formulations in crop production: An overview. International Journal of Agricultural Sciences and Veterinary Medicine, 8(2), 38–46. [Google Scholar]
21. Chen, Z. C., Peng, W. T., Li, J., Liao, H. (2018). Functional dissection and transport mechanism of magnesium in plants. Seminars in Cell & Developmental Biology, 74, 142–152. https://doi.org/10.1016/j.semcdb.2017.08.005 [Google Scholar]
22. Hauer-Jákli, M., Tränkner, M. (2019). Critical leaf magnesium thresholds and the impact of magnesium on plant growth and photo-oxidative defence: A systematic review and meta-analysis from 70 years of research. Frontiers in Plant Science, 10, 766. https://doi.org/10.3389/fpls.2019.00766 [Google Scholar] [PubMed] [CrossRef]
23. Yan, B., Hou, Y. (2018). Effect of soil magnesium on plants: A review. IOP Conference Series: Earth and Environmental Science, 170, 022168. https://doi.org/10.1088/1755-1315/170/2/022168 [Google Scholar] [CrossRef]
24. Marschner, H. (2012). Marschner’s mineral nutrition of higher plants. New York: Academic Press. [Google Scholar]
25. Gout, E., Rébeillé, F., Douce, R., Bligny, R. (2014). Interplay of Mg2+, ADP, and ATP in the cytosol and mitochondria: Unravelling the role of Mg2+ in cell respiration. Proceedings of the National Academy of Sciences of the United States of America, 111(43), E4560–E4567. https://doi.org/10.1073/pnas.1406251111 [Google Scholar] [PubMed] [CrossRef]
26. Summart, J., Thanonkeo, P., Panichajakul, S., Prathepha, P., McManus, M. T. (2010). Effect of salt stress on growth, inorganic ion and proline accumulation in Thai aromatic rice, Khao Dawk Mali 105, callus culture. African Journal Biotechnology, 9(2), 145–152. [Google Scholar]
27. Duval, F. D., Renard, M., Jaquinod, M., Biou, V., Montrichard, F. et al. (2002). Differential expression and functional analysis of three calmodulin isoforms in germinating pea (Pisum sativum L.) seeds. Plant Journal, 32(4), 481–493. https://doi.org/10.1046/j.1365-313X.2002.01409.x [Google Scholar] [PubMed] [CrossRef]
28. Clapham, D. E. (2007). Calcium signaling. Cell, 131(6), 1047–1058. https://doi.org/10.1016/j.cell.2007.11.028 [Google Scholar] [PubMed] [CrossRef]
29. Khan, S., Verma, G., Sharma, S. (2010). A novel Ca2+ activated protease from germinating Vigna radiata seeds and its role in storage protein mobilization. Journal of Plant Physiology, 167(11), 855–861. https://doi.org/10.1016/j.jplph.2010.01.010 [Google Scholar] [PubMed] [CrossRef]
30. Verma, G., Khan, S., Agarwal, S. K., Sharma, S. (2019). Role of apoplastic calcium during germination and initial stages of seedling establishment in Vigna radiata seeds. Journal of Plant Physiology, 236, 66–73. https://doi.org/10.1016/j.jplph.2019.02.009 [Google Scholar] [PubMed] [CrossRef]
31. Imran, M., Mahmood, A., Neumann, G., Boelt, B. (2021). Zinc seed priming improves spinach germination at low temperature. Agriculture, 11(3), 271. https://doi.org/10.3390/agriculture11030271 [Google Scholar] [CrossRef]
32. Genc, Y., McDonald, G. K., Graham, R. D. (2005). The interactive effects of zinc and salt on growth of wheat. Plant nutrition for food security, human health and environmental protection, pp. 548–549. Beijing, China: Tsinghua University Press. [Google Scholar]
33. Colmenero-Flores, J. M., Franco-Navarro, J. D., Cubero-Font, P., Peinado-Torrubia, P., Rosales, M. A. (2019). Chloride as a beneficial macronutrient in higher plants: New roles and regulation. International Journal of Molecular Sciences, 20(19), 4686. https://doi.org/10.3390/ijms20194686 [Google Scholar] [PubMed] [CrossRef]
34. Day, S. (2022). Impact of seed priming on germination performance of fresh and aged seeds of canola. International Journal of Agriculture Environment and Food Sciences, 6(1), 37–40. https://doi.org/10.31015/jaefs.2022.1.6 [Google Scholar] [CrossRef]
35. ISTA (2017). International rules for seed testing. Basserdorf, Switzerland: International Seed Testing Association. [Google Scholar]
36. Day, S. (2016). Determining the impact of excessive boron on some growth characters and some nutrients at the early growth stage of sunflower (Helianthus annuus L.). Fresenius Environmental Bulletin, 25(10), 4294–4298. [Google Scholar]
37. Day, S., Çıkılı, Y., Aasim, M. (2017). Screening of three safflower (Carthamus tinctorius L.) cultivars under boron stress. Acta Scientarum Polonorum Hortorum Cultus, 16(5), 109–116. https://doi.org/10.24326/asphc.2017.5.11 [Google Scholar] [CrossRef]
38. Mielezrski, F., Marcos-Filho, J. (2013). Assessment of physiological potential of stored pea (Pisum sativum L.) seeds. Journal of Seed Science, 35(1), 42–50. [Google Scholar]
39. Hoekstra, F. A., Golovina, E. A., Buitink, J. (2001). Mechanisms of plant desiccation tolerance. Trends in Plant Science, 6(9), 431–438. https://doi.org/10.1016/S1360-1385(01)02052-0 [Google Scholar] [PubMed] [CrossRef]
40. Vertucci, C. W., Farrant, J. M. (1995). Acquisition and loss of desiccation tolerance. In: Seed development and germination, pp. 237–271. Newyork: Marcel Dekker Inc. [Google Scholar]
41. Ceylan, Y., Kutman, U. B., Mengutay, M., Cakmak, I. (2016). Magnesium applications to growth medium and foliage affect the starch distribution, increase the grain size and improve the seed germination in wheat. Plant and Soil, 406(1–2), 145–156. https://doi.org/10.1007/s11104-016-2871-8 [Google Scholar] [CrossRef]
42. Zhang, B., Cakmak, I., Feng, J., Yu, C., Chen, X. et al. (2020). Magnesium deficiency reduced the yield and seed germination in wax gourd by affecting the carbohydrate translocation. Frontiers in Plant Science, 11, 797. https://doi.org/10.3389/fpls.2020.00797 [Google Scholar] [PubMed] [CrossRef]
43. Bhowmick, M. K. (2018). Seed priming: A low-cost technology for resource-poor farmers in improving pulse productivity. In: Advances in seed priming, pp. 187–208. Singapore: Springer. https://doi.org/10.1007/978-981-13-0032-5_11 [Google Scholar] [CrossRef]
44. Pradeepkumar, T., Peter, K. V., Jose, M. A. (2020). Innovations in breaking the yield barrier of tropical vegetables. International Journal of Innovative Horticulture, 9(2), 120–130. https://doi.org/10.5958/2582-2527.2020.00015.9 [Google Scholar] [CrossRef]
45. Stanković, M., Topuzović, M., Marković, A., Pavlović, D., Đelić, G. et al. (2010). Influence of zinc (Zn) on germination of wheat (Triticum aestivum L.). Biotechnology & Biotechnological Equipment, 24(2), 236–239. https://doi.org/10.1080/13102818.2010.10817842 [Google Scholar] [CrossRef]
46. Lutts, S., Benincasa, P., Wojtyla, L., Kubala, S. S., Pace, R. et al. (2016). Seed priming: New comprehensive approaches for an old empirical technique. In: Araujo, S., Balestrazzi, A. (Eds.New challenges in seed biology-basic and translational research driving seed technology. London: IntechOpen. https://doi.org/10.5772/64420 [Google Scholar] [CrossRef]
47. Brooks, S., Athinuwat, D., Chiangmai, P. N. (2020). Enhancing germination and seedling vigor of upland rice seed under salinity and water stresses by osmopriming. Science & Technology Asia, 25(2), 63–74. https://doi.org/10.14456/scitechasia.2020.21 [Google Scholar] [CrossRef]
48. Wang, X., Dingxuan, Q., Shi, M. (2021). Calcium amendment for improved germination, plant growth, and leaf photosynthetic electron transport in oat (Avena sativa) under NaCl stress. PLoS One, 16(8), e0256529. https://doi.org/10.1371/journal.pone.0256529 [Google Scholar] [PubMed] [CrossRef]
49. Fatonah, K., Suliansyah, I., Rozen, N. (2017). Electrical conductivity for seed vigor test in sorghum (Sorghum bicolor). Cell Biology and Development, 1(1), 6–12. https://doi.org/10.13057/cellbioldev/v010102 [Google Scholar] [CrossRef]
50. Szemruch, C., Gallo, C., Murcia, M., Esquivel, M., García, F. et al. (2019). Electrical conductivity test for predict sunflower seeds vigor. SSRG International Journal of Agriculture & Environmental Science (SSRG-IJAES), 6(4), 118–127. [Google Scholar]
51. Ashraf, M., Foolad, M. R. (2005). Pre-sowing seed treatment: A shotgun approach to improve germination growth and crop yield under saline and non-saline conditions. Advances in Agronomy, 88, 223–271. https://doi.org/10.1016/S0065-2113(05)88006-X [Google Scholar] [CrossRef]
52. Kubala, S., Garnczarska, M., Wojtyla, Ł., Clippe, A., Kosmala, A. et al. (2015). Deciphering priming-induced improvement of rapeseed (Brassica napus L.) germination through an integrated transcriptomic and proteomic approach. Plant Science, 231, 94–113. https://doi.org/10.1016/j.plantsci.2014.11.008 [Google Scholar] [PubMed] [CrossRef]
53. Ashraf, M. A., Iqbal, M., Rasheed, R., Hussain, I., Perveen, S. et al. (2018). Dynamic proline metabolism: Importance and regulation in water-limited environments. In: Plant metabolites and regulation under environmental stress, pp. 323–336. Cambridge: Academic Press. https://doi.org/10.1016/B978-0-12-812689-9.00016-9 [Google Scholar] [CrossRef]
Cite This Article
 Copyright © 2023 The Author(s). Published by Tech Science Press.
Copyright © 2023 The Author(s). Published by Tech Science Press.This work is licensed under a Creative Commons Attribution 4.0 International License , which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
 Downloads
Downloads
 Citation Tools
Citation Tools