
 Submit a Paper
Submit a Paper Propose a Special lssue
Propose a Special lssue Open Access
Open Access
REVIEW

Gut Microbiota-Derived Exosomes Are Unique Natural Nanocarriers for Therapeutics
1 Department of Clinical Pharmacology, Faculty of Medicine, King Abdulaziz University, Jeddah, Saudi Arabia
2 Department of Pharmaceutics, Faculty of Pharmacy, King Abdulaziz University, Jeddah, Saudi Arabia
* Corresponding Author: Shadab Md. Email:
(This article belongs to the Special Issue: Bioactive Natural Components as Regulators of Cellular Pathways and Disease Progression)
BIOCELL 2026, 50(7), 3 https://doi.org/10.32604/biocell.2026.077548
Received 11 December 2025; Accepted 25 February 2026; Issue published 29 June 2026
 View Full Text
View Full Text Download PDF
Download PDFAbstract
Gut microbiota-derived exosomes (MDEs) have emerged as a novel class of drug delivery and are secreted by bacteria, fungi, and archaea in the human microbiota within the human intestinal ecosystem and possess inherent biocompatibility and lower immunogenicity, enabling seamless integration within host intestinal and systemic bioenvironments. This review elucidates the cellular and molecular mechanisms governing MDE function, explaining how their unique lipid bilayer composition facilitates cellular entry via receptor-mediated endocytosis and membrane fusion. This review discusses how gut MDEs traverse biological barriers, such as the blood-brain barrier and intestinal mucosa, by modulating tight junction proteins and accurately transporting cargoes to desired tissues. Upon internalization, MDE cargoes actively modulate intracellular signaling cascades. Various drugs, including RNA, proteins, and small molecules, can be loaded into MDEs via physical or biological methods. Furthermore, bioengineering strategies to functionalize MDE surfaces with specific ligands for precise molecular targeting are evaluated. While obstacles regarding standardized production and quality control remain, gut MDEs have great promise in achieving personalized and precision medicine by targeting diseases such as cancer, inflammatory diseases, and male infertility. Future clinical translation relies on exploring these molecular interactions to generate highly efficient, engineered nanocarriers.Graphic Abstract
Keywords
Exosomes, small vesicles with diameters from 30 to 150 nm, play an important role in intercellular communication and have promising applications in drug delivery, compared with traditional carriers [1,2]. In comparison to traditional synthetic carriers, exosomes exhibit significantly lower toxicity and higher biological tolerance, while their unique structural integrity effectively protects therapeutic cargo from enzymatic and systemic degradation [1,3]. Fig. 1 illustrates the heterogeneous origins of exosomes, including microbiota-derived exosomes (from gut bacteria, fungi, and bacteriophage), mesenchymal stem cell-derived exosomes, tumor-derived exosomes, immune cell-derived exosomes, and neuronal cell-derived exosomes. The fate of the exosomes is also indicated.

Figure 1: Heterogeneous origins of exosomes and their fate. This original figure is prepared with Mind the Graph (www.mindthegraph.com) and modified using https://BioRender.com.
Microbiota-derived exosomes (MDEs) have recently emerged as a promising therapeutic approach for the treatment of diseases, including neurological diseases, by harnessing the advantages of exosome delivery systems and the interaction within the gut-brain axis [2,4]. In this review, outer membrane vesicles (OMVs) released by Gram-negative bacteria or simply membrane vesicles (MVs) by Gram-positive bacteria are also indicated by MDEs. For example, MDEs from Bacteroides fragilis containing Polysaccharide A (PSA) have been reported [5]. Beyond their local roles, gut-derived MDEs are currently being evaluated as a novel therapeutic paradigm for Alzheimer’s disease (AD) because gut bacteria significantly impact neurodegenerative development, and their modification has been shown to enhance the efficacy of treatments utilizing stem cell-derived exosomes [4]. A generalized comparison of MDEs with exosomes derived from different origins is provided in Table 1.
Table 1: Comparison of MDEs with exosomes derived from different origins.
| Aspect | Microbiota-Derived Exosomes [6,7,8] | Mesenchymal Stem Cell-Derived Exosomes [9,10,11] | Tumor-Derived Exosomes [12,13] | Immune Cell-Derived Exosomes [14,15] | Neural Cell-Derived Exosomes [16,17] |
|---|---|---|---|---|---|
| Origin | Secreted by bacteria, fungi, and archaea in the gut microbiome or other microbial ecosystems | Secreted by mesenchymal stem cells (bone marrow, adipose, etc.) | Secreted by cancer cells from tumors | Secreted by immune cells (macrophages, T cells, etc.) | Secreted by neurons and neural progenitor cells |
| Main cargo | Microbial DNA, RNA, proteins, and metabolites | mRNA, microRNA, proteins, growth factors | Tumor antigens, microRNA, oncogenic proteins | Cytokines, chemokines, antigenic peptides, microRNAs | Neuronal proteins, microRNAs, and lipids |
| Biological function | Regulate host-microbiome interactions, gut homeostasis, and immune modulation | Tissue repair, immunomodulation, and anti-inflammatory effects | Promote tumor growth, metastasis, and immune evasion | Immune surveillance, antigen presentation, and immune modulation | Neuronal communication, neuroprotection, and synaptic plasticity |
| Therapeutic potential | Gut health, metabolic diseases, and immune modulation | Regenerative medicine, anti-inflammatory, tissue repair | Cancer immunotherapy, targeting the tumor microenvironment | Cancer immunotherapy, autoimmune diseases | Neurological disorders (Alzheimer’s, Parkinson’s, etc.) |
| Immunomodulatory effect | Diverse: pro- or anti-inflammatory depending on microbial origin | Anti-inflammatory, immune cell regulation | Immunosuppressive, immune evasion | Modulate immune responses, promote T-cell activation | Immune regulation in neuroinflammatory diseases |
| Cellular uptake mechanism | Primarily via endocytosis or phagocytosis by intestinal cells | Uptaken by macrophages, fibroblasts, and endothelial cells | Taken up by immune and tumor cells | Uptaken by antigen-presenting cells, T cells | Uptaken by neurons, glial cells |
| Influence on disease | Modulate gut-brain axis, inflammation, metabolic disorders | Promote wound healing, reduce inflammation, and regenerate tissues | Promote tumor progression, metastasis, and drug resistance | Modulate autoimmune diseases, tumor response | Implicated in neurodegeneration, neuroinflammation |
| Stability in circulation | Relatively high due to bacterial membrane composition | Stable in body fluids like blood and CSF | High stability due to cancer cell adaptations | High stability to enhance immune signaling | Moderate stability, affected by neurodegenerative conditions |
| Interaction with host cells | Interact with intestinal epithelial cells, immune cells | Interact with immune cells, fibroblasts, and endothelial cells | Direct interaction with tumor stroma and immune cells | Interact with immune cells, particularly T cells | Interact with neurons, glial cells |
| Role in drug delivery | Not widely explored, but potential in gut health | Widely studied for drug delivery in tissue regeneration and inflammation | Being investigated for cancer drug delivery | Potential in delivering immune-modulatory drugs | Potential for CNS drug delivery |
| Effect on inflammation | Can either promote or suppress inflammation depending on the microbial source | Anti-inflammatory, reduces cytokine storms | Promote tumor-associated inflammation | Anti-inflammatory or pro-inflammatory, depending on cell type | Modulate neuroinflammation, potentially neuroprotective |
| Tumor-related role | Some microbiome exosomes may influence cancer progression | Anti-cancer effects in some models promote immune surveillance | Pro-tumorigenic, promote metastasis, and drug resistance | Can promote or inhibit tumor progression | Rarely involved, but neuroimmune functions may impact gliomas |
| Challenges in application | Isolation and characterization due to diverse microbiome sources | Large-scale production and isolation, maintaining therapeutic consistency | Safety concerns due to tumor origin | Modulation of immune responses without off-target effects | Blood-brain barrier crossing, targeted delivery |
| Example of usage | Microbiota transplants or probiotics containing exosomes | Treatment of osteoarthritis, wound healing, and myocardial infarction | Cancer vaccine delivery, chemotherapy resistance reduction | Cancer vaccines, autoimmune disease treatment | Neuroprotective exosomes for Alzheimer’s and Parkinson’s |
The present review focuses on the therapeutic potential of MDEs. However, where specific data on MDEs is limited, this review draws upon parallel mechanisms observed in synthetic and mammalian exosome systems. These established models provide the theoretical and practical framework necessary to understand how MDEs can be engineered and utilized for drug delivery.
2 Overview of Gut Microbiota and Its Role in Human Health
The human gastrointestinal tract contains trillions of bacteria and viruses that form complex ecosystems on our skin and the tracts of our internal organs [18]. The human gastrointestinal tract hosts a complex system where the gut contains four million different types of bacteria, representing a microbial population that is approximately ten times greater than the total number of human cells within the host [19,20]. They provide numerous health benefits by providing nutrients and assisting the body with metabolizing food, thereby protecting the intestinal barrier [21,22]. Functioning as a critical regulatory hub, the gut modulates the immune system while facilitating a complex interaction between the gut and the brain that profoundly helps to influence host mood and behavior [20]. The individual genetics and diets of people play a role in determining the functional capacity of the microorganisms [20,23,24]. The mucosal immune system is a specialized barrier distributed across the intestinal lining to protect the integrity of the gastrointestinal tract. The interaction of gut flora with T cells, the core cells of adaptive immunity, is a primary factor in determining the host immune response. Dybiosis is a consequence of an overabundance of pathogens within the gut and leads to adverse health conditions [25]. Therefore, knowledge of how gut-resident microorganisms function will enable the design of therapies to maintain healthy microflora [19,25].
The microbiota serve as a guide to the host immune system to help it in its efforts of surveillance. This important role of the microbiota allows for the management of infections and transforms the host and the microbes into symbiotic partners [26]. An ecosystem of one hundred trillion microbes greatly impacts the overall health of the human host [27]. The gut microbiota and the mucosal immune system maintain a vital bidirectional relationship. The microbiota facilitates the maturation of the mucosal immune system, which in turn governs the spatial distribution of gut bacteria [28]. Functioning as a barrier throughout the gastrointestinal tract, the mucosal immune system ensures intestinal integrity and discriminates between commensal symbionts and pathogenic threats. This regulatory capacity relies on the crosstalk between gut flora and T cells, a process that is fundamental to adaptive immunity. These microbial interactions directly inform the generation of antibodies and memory immune cells, establishing the foundation for the host’s long-term immunological memory [28,29,30]. Disruptions to this microbial balance can trigger immune dysfunctions, increasing the host’s vulnerability to various inflammatory conditions [19,23]. Therefore, a deeper investigation into these mechanistic pathways is required to advance therapeutic strategies for immune-mediated diseases [31,32].
MDEs interact with a broad range of innate and adaptive immune cells. Their molecular cargo largely determines whether they cause pro- or anti-inflammatory responses [33,34]. At mucosal surfaces, MDEs are taken up by macrophages and dendritic cells via endocytosis and engagement of pattern-recognition receptors, including Toll-like receptors. This uptake triggers context-dependent production of cytokines such as TNF-α, IL-6, IL-12, IL-1β, and IL-10, thereby shaping downstream immune responses [34,35]. Vesicles from pathogenic bacteria often carry lipoproteins, lipopolysaccharide, and other pathogen-associated molecular patterns that activate NF-κB signalling and favour Th1- and Th17-biased responses. In contrast, commensal-derived MDEs can promote tolerogenic dendritic cells and regulatory T cells, limiting excessive inflammation and supporting gut immune homeostasis [33,35,36]. EVs produced by the gut commensal Bacteroides thetaiotaomicron induce an anti-inflammatory programme in bone-marrow-derived macrophages. These macrophages display increased IL-10 production and reduced responsiveness to secondary inflammatory stimuli, consistent with features of innate immune tolerance [37]. By transferring bacterial antigens, nucleic acids, and metabolites, MDEs also modulate antigen presentation and shape CD4+ T-cell differentiation. They further influence B-cell antibody production, thereby linking specific microbiota signatures to systemic immune phenotypes [34,38]. The available data indicate that MDEs function as potent immunological messengers. Their effects on immune cell activity depend on both microbial origin and cargo composition in physiological and pathological contexts [33,36,39].
3 Functional Role of MDEs in Disease Progression and Their Therapeutic Potential
MDEs play an essential role in intercellular communication and communication between different biological kingdoms, and are responsible for maintaining balance in the gut and affecting other physical systems [32]. Intercellular and inter-kingdom forms of communication occur through extracellular vesicles (EVs) that facilitate the transfer of the biologically active components (or bioactive components) across the appropriate biological barriers [40,41]. These components, also known as cargos, include proteins, lipids, and RNA, but this list is not exhaustive. MDEs regulate the microbiota-gut-brain network through the delivery of neuroactive components to the CNS; thus, they may provide a possible therapeutic benefit in treating neuropathologies and mood disorders [41]. MDEs maintain performance in the gut and modulate immune systems as key determinants of host-microbe relationships and have great potential therapeutic activity against pathologies, including inflammatory bowel disease (IBD), colorectal cancer (CRC), and neurological disorders [40,42].
The gut microbiome is crucial to health, but can also contribute to disease [43,44]. In particular, two of the most destructive diseases affecting the gut today (and even the body itself for some) are IBD and CRC. IBD is characterized, to a degree, by chronic inflammation of the gastrointestinal (GI) tract and shifting of the local gut microbial community. People with IBD are likely to have a very different microbiome composition compared to healthy controls [45]. This can lead to the disruption of the mucosal barrier and continual activation of inflammation. Bacteria, such as Fusobacterium nucleatum, Escherichia coli, and enterotoxigenic Bacteroides fragilis, are linked with colorectal cancer [44]. Established knowledge is that gut bacteria can impact IBD and related diseases by influencing the immune system [46]. Next-generation organoid microfluidics technology may advance understanding of IBD-associated microbial-epithelial interactions and develop new microbial therapies for the condition [47]. Having an approach that concentrates on an at-risk individual’s gut bacteria may allow for better disease outcomes and general lifestyle improvement [44].
Biogenic exosomes secreted from gut bacteria demonstrate potential in the treatment of metabolic issues and malignancies such as cancer by functioning as specialized nanoparticles that alter and reprogram the host immune system. In order to improve their therapeutic potential, these vesicles can be bio-engineered, complementing findings that exosomes produced from immune cells exhibit the unique potential for eliciting potent antitumor immune responses. For better drug delivery, technology has improved exosome modification. Therapeutic cargoes are transported to particular tumor sites by engineered exosomes [48]. These vesicles have strong anti-tumor properties. The effectiveness of immunotherapy and cancer treatment is influenced by gut microbes. Response to immune checkpoint inhibitor therapy is determined by the interaction between the immune system and microbiome. In order to improve the response to cancer therapy, methods such as fecal microbiota transplantation are being investigated [49]. In models of metabolic stress, these exosomes successfully restored β-cell function and mass by modulating inflammatory pathways, specifically targeting the NLRP3 inflammasome [50].
The ability to engineer exosomes to improve their functions and target specific sites makes them versatile tools for novel therapies. Developing therapeutic strategies that target the gut microbiome to improve patient outcomes would be highly beneficial [51]. Fig. 2 illustrates three therapeutic modalities of using MDEs: inflammatory bowel diseases, cancer chemotherapy, and CNS disorders, crossing the blood-brain barrier (BBB).

Figure 2: Therapeutic applications of microbiota-derived exosomes (MDEs). Three therapeutic modalities of using MDEs are presented here. MDEs have the potential to advance cancer therapy by targeting tumor cells, carrying anticancer drugs, or improving immune functions. The inhibition of gut inflammation and promotion of gut tissue repair are proposed as the other two therapeutic targets in inflammatory bowel disease (IBD) therapy. The application of microbiota-derived exosomes can also promote the delivery of therapeutic reagents to the brain by crossing the BBB, which offers hope for the treatment of CNS diseases. This original figure is prepared with Mind the Graph (www.mindthegraph.com) and modified using https://BioRender.com.
4 Advantages of MDEs in Drug Delivery
Therapies with MDEs are a strategy that hybridizes the inherent advantages of synthetic nanoparticles and cell-based vehicles, as well as reduces their inherent shortcomings [52]. MDEs are endogenous extracellular vesicles that are enclosed by a bilayer lipid membrane and have a capacity to transport diverse biomolecules, including proteins and nucleic acids [53]. When utilized as a therapeutic tool, MDEs offer a distinct clinical advantage because they are inherently nontoxic and nonimmunogenic, minimizing the risk of adverse inflammatory reactions [3,53]. They can also carry various cargos within a stable membrane, which protects them from degradation and facilitates transfer into recipient cells [40]. MDEs can be genetically engineered to express specific nucleic acids or proteins and to carry chosen targeting molecules on their surfaces, resulting in highly specific cell targeting [52,54]. While historically most work has been focused on mammalian cell-derived exosomes, the unique biochemical profile of microbiota-derived vesicles is now receiving increased attention for its specific targeting capabilities. Plant-derived exosomes (PDEs) are also being studied because they have a significant cancer chemopreventive effect with minimal toxicity, unlike the undesirable effects of other drug delivery methods [55]. There are several different exosome sources for drug delivery. MDEs represent a potential versatile drug delivery vehicle that is biocompatible, conducive to targeted delivery, and caters to cargo protection, and hence their applications are diverse, from cancer management to therapy of neurodegenerative disorders [52]. Nevertheless, issues pertaining to fabrication standardization, approaches of drug-loading, and long-term safety assessment for clinical use prevail [3,56].
4.1 Biocompatibility and Low Immunogenicity
To date, exosomes derived from immune cells have been utilized as versatile drug delivery systems for cancer therapy, mainly because of their good biocompatibility and low immune responses [57]. As bioactive particles secreted by diverse cells under various physiological or pathological conditions, exosomes play an important role in cell-to-cell communication, with their well-characterized immunomodulatory functions. Although exosomes originating from immune cells are generally characterized by low immunogenicity, the gut microbiota is indispensable for the development and proper functioning of the host immune system. This principle is substantiated by evidence from germ-free animal models, which demonstrate significant immune system deficiencies and heightened susceptibility to both infectious and autoimmune disease [58]. These findings highlight a complex interplay: while individual immune cell-derived exosomes are weak immune activators, the collective microbiota profoundly influences host immune competence. It is precisely this low immunogenicity, combined with high biocompatibility, that has led to the extensive investigation of these exosomes as delivery vehicles for anticancer therapeutics. However, the bidirectional effects of such exosomes on the host immune system within the context of varying gut microbial compositions remain to be fully elucidated.
MDEs offer a number of advantages as a drug delivery vehicle for treating intestinal diseases compared to synthetic nanocarriers. They are characterized by high biocompatibility with low immunogenicity compared to synthetic nanoparticle carriers [59], and integrate seamlessly into natural cellular systems without triggering rejection. Mechanistically, this low immunogenicity is often attributed to the lack of specific immunogenic surface markers found on synthetic carriers and the presence of ‘don’t eat me’ signals that prevent clearance by the reticuloendothelial system. Due to the fundamental structural homology between exosome membranes and the lipid bilayer of host cell membranes, recipient cells exhibit a significantly higher degree of receptivity toward these biogenic vesicles, which facilitates more efficient internalization compared to traditional synthetic delivery systems. Therefore, exosome lipids enhance their biocompatibility and increase the likelihood of them being taken up by acceptor cells [60,61]. Researchers believe that plant-derived exosome represents a potential microorganism-free oral vehicle for treating various gut diseases [59]. The special affinity between exosomes and gut flora, therefore, makes MDEs another type of cell-free natural carrier for the treatment of gastrointestinal diseases. Owing to their naturally derived origin, these vesicles provide significant therapeutic benefits by maintaining high compatibility with complex biological systems and possessing a remarkably low potential for triggering adverse systemic immune responses. Because of their exceptional ability to traverse biological barriers and deliver therapeutic cargo, MDEs are superior to synthetic nanocarriers in targeted drug delivery.
Gut-derived MDEs possess an inherent biocompatibility that serves to naturally enable complex, inter-kingdom cell communication pathways essential for maintaining host homeostasis. Plant-derived exosomes particularly have the capability to crosstalk with the gut microbiome as well as the host physiology. Plant-derived exosomes exhibit a marked biocompatibility due to their specific uptake pathways by gut commensals that eventually ameliorate the barrier function of the host without adverse immune activation [62]. Their utility as drug delivery systems and diagnostic biomarkers underscores their importance going forward.
4.2 Targeting Abilities and Bioavailability
Exosomes derived from gut microbiota are also excellent candidates for specific tissue targeting and targeted delivery of therapeutic cargo. Within the gut-liver axis, microbial-derived MDEs operate as significant modulators with dual targeting to the intestinal epithelium and hepatic tissues. Gut-derived MDEs function as specialized conduits that spread diverse bioactive payloads throughout the host architecture, thereby directly modulating essential aspects of host metabolism and systemic immune responses. Bacterial sphingolipids derived from Bacteroides thetaiotaomicron traffic from the gut to colon and liver in a murine experimental model [63]. This provides evidence that MDE cargo delivered to the liver has a direct impact on hepatic function and metabolism. Communication is already bidirectional as compounds produced in the liver modify gut microbiota composition with direct implications for liver health [64]. The potential of MDEs in focused interventions is highlighted by this complex interaction. Exosomes transport bioactive substances to both distant and local tissues in order to alter host physiology [65]. The elucidation of this mechanism increases the probability that a wide range of chronic metabolic diseases can be effectively treated using microbiome-based strategies centered on exosomal trafficking. Additionally, through the “gut-testis” axis, gut MDEs can affect distant organs. They alter miRNA expression in exosomes in circulation and hinder with spermatogenesis in the testes. The effects of circulating exosomes on mouse spermatogenesis are well known [66].
Because of their great biocompatibility and circulatory stability, exosomes are promising drug delivery vehicles [67,68]. The elucidation of this mechanism substantially increases the probability that a wide range of chronic metabolic diseases can be effectively treated using innovative microbiome-based strategies centered on exosomal trafficking. A vast array of complex physiological processes, extending from local intestinal homeostasis to distant organ regulation, are dynamically regulated by the specialized cargo carried within MDEs. The dysbiosis of the gut microbiota affects murine spermatogenesis and modifies the expression of miRNA in circulating exosomes [66]. However, there are still no specific studies on MDEs for improving systemic bioavailability. However, exosomes are more stable and soluble in substances like curcumin [69]. Delivery methods based on exosomes appear promising for enhancing circulation time and getting past biological barriers [70]. Thus, MDEs offer a promising avenue for further investigation into the stability and bioavailability of drugs.
4.3 Crossing Biological Barriers
More generally, MDEs are essential for passage through biological barriers such as the blood-brain barrier (BBB) and intestinal mucosa, thus contributing to the communication of gut microbiota with peripheral organs. Gut-derived MDEs and related biogenic nanoparticles effectively navigate along cellular corridors by exploiting endogenous pathways of cellular communication and vesicular transport [71]. They cross barriers more efficiently than other systems because their payload and bioavailability are biologically predisposed [72]. While various synthetic nanoparticle systems share some of these physicochemical properties, microbiota-derived exosomes excel as therapeutic vehicles due to their unique and evolutionary role in facilitating seamless cellular communication. The structural integrity and selective permeability of critical interfaces, such as the BBB and the intestinal mucosa, are profoundly influenced by the metabolic activity and circulating factors derived from the gut microbiota. Exchange of gut microbiota-derived circulating factors may lead to modification of the BBB, facilitating passage of exosomes and molecules into the brain [73]. This passage is likely mediated by transcytosis, where MDEs are encapsulated in vesicles on the luminal side, transported across the cytoplasm, and released on the abluminal side. In addition, the gut microbiota regulates the permeability of these barriers, creating a favorable environment that promotes the efficient passage of MDEs from the gut lumen into systemic circulation [74,75]. Thus, MDEs are particularly adept at crossing these barriers on account of their intrinsic properties and the regulatory capacity of gut microbiota over these barriers. The ability to traverse tight junctions and modulate the physiology of these barriers allows efficient crossing of these interfaces from one compartment to another. Elucidation of the molecular pathways and biological signaling mechanisms will provide novel therapeutic targets for different diseases [71,72].
It has been demonstrated that a new treatment for ischemic stroke is possible by using bacteria-derived MDEs to deliver the drug pioglitazone directly to the brain. These MDEs are designed to be taken up by immune cells called neutrophils, which naturally travel to the site of brain injury. This “hitchhiking” allows the drug to bypass the blood-brain barrier and reach the damaged brain tissue [76]. Furthermore, certain bacterial MDEs have been observed to bypass the BBB and localize in brain tissue, although this can sometimes lead to pathogenic activation of glial cells [77].
5 Cellular and Molecular Mechanisms of MDE Action
Understanding the molecular mechanisms behind cellular entry and intracellular modulation is essential to utilizing MDEs’ therapeutic potential. Through active interactions with host cell membranes and signaling pathways, these entities perform functions beyond those of passive drug carriers (Fig. 3).
Similar to mammalian cells, the lipid bilayer compositions of MDEs facilitate the uptake of MDEs as a function of the enhanced biocompatibility of exosome lipids, which increases the chances of MDEs being taken up by acceptor cells [60,61]. Host cells typically utilize receptor-mediated endocytosis and membrane fusion to internalize MDEs. Ginger-derived exosome-like nanoparticles (GELNs) are internalized by specific gut commensals (Lactobacillaceae) via a ligand-receptor lipid-dependent mechanism [62]. Once at the target site or within the target cell, the interaction of the respective surface structures allows uptake of the MDE. This mechanism is especially critical for crossing biological barriers. The presence of the gut microbiota has been shown to modulate the transport of exosomes across intestinal epithelial cells by modulating tight junction protein expression, facilitating the passage of exosomes [75]. Additionally, microbiota-derived short-chain fatty acids stemming from commensal bacteria can modulate the structure of the cell in order to facilitate exosomes’ passage across biological barriers [74].
5.1.1 Sub-Pathways of Receptor-Mediated Endocytosis
Receptor-mediated endocytosis of MDEs and bacterial EVs (BEVs) often uses clathrin-mediated endocytosis. This pathway depends on AP-2 adaptors and dynamin for vesicle scission. It is reported for Gram-negative OMVs such as those from Escherichia coli that enter intestinal epithelial cells, including HT-29, and for enterohemorrhagic Escherichia coli EVs. Caveolae-mediated endocytosis involves caveolin-1 and lipid rafts. It supports the uptake of Helicobacter pylori EVs. Macropinocytosis supports non-specific fluid-phase uptake of larger bacterial EVs, especially in immune cells. Phagocytosis is a major uptake route in macrophages for Gram-negative OMVs that display pathogen-associated molecular patterns. These uptake pathways deliver EVs into endosomal compartments. A fraction of vesicles can then escape from endosomes and release their cargo into the cytosol [35,78,79].
5.1.2 Role of Microbial Lipids in Membrane Fusion
Acidic pH and electrolyte conditions diminish electrostatic repulsion between Gram-negative OMVs, thereby promoting their aggregation and fusion. Escherichia coli OMVs exhibit robust lipid mixing under low pH environments. Lipopolysaccharide (LPS) truncation, as observed in modified Escherichia coli (ClearColi) OMVs, lowers the apparent isoelectric point and alters the critical coagulation concentration, facilitating vesicle aggregation and fusion under saline conditions where full-length LPS-bearing Escherichia coli OMVs demonstrate reduced fusogenicity. This indicates that intact LPS imposes a steric and electrostatic barrier to salt-induced lipid mixing. Divalent cations, including Ca2+ and Mg2+, mediate crosslinking of LPS and phospholipids, which stabilizes the outer membrane structure and restricts lateral membrane fluidity, thereby constraining OMV fusion despite the fusogenic influence of acidity. These observations identified key physicochemical parameters that govern OMV fusion in vitro but do not provide direct assessments of OMV-host membrane fusion, endocytic evasion, or cytosolic cargo dissemination [80].
5.1.3 Short-Chain Fatty Acids in Facilitating Transcytosis
Short-chain fatty acids (SCFAs), such as acetate, propionate, and butyrate, are produced by commensal microbiota. These SCFAs signal through FFAR2/GPR43 and FFAR3/GPR41 receptors on intestinal epithelial and immune cells. They upregulate tight junction proteins, including ZO-1 and occludin, to support barrier integrity. Butyrate acts as a histone deacetylase inhibitor. This action leads to epigenetic regulation of gene expression in epithelial and immune cells. The result supports mucosal barrier function, gut motility, and reduced inflammation [81,82].
5.2 Intracellular Signaling and Modulation
Upon internalization, MDEs release their bioactive cargo comprising miRNAs, proteins, and lipids into the cytoplasm, where they modulate critical signaling cascades.
Pathogenic MDEs activate TLR4/NF-κB via lipopolysaccharides, while therapeutic MDEs inhibit NLRP3 inflammasome. There are many types of inflammatory situations in which MDEs can take on a ‘double-edged sword’ role. The Toll-like receptor 4 (TLR4)/Nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) signalling cascade may be activated by pathogenic MDEs. In fact, it is through that route by which vesicles carrying gut bacterial lipopolysaccharides drive inflammation [83]. In order to elicit the release of neurotoxic factors, MDEs can activate astrocytes through inflammatory markers. Such activation of cells is entirely dependent on the NF-κB protein [77].
Therapeutic MDEs seek to actively inhibit those pathways, however, allowing inflammation to be effectively resolved. Newer work has suggested that MDEs may directly target the NLRP3 inflammasome of the innate immune system. Repairing pancreatic β-cell function has been enhanced by gut-derived exosomes rich in angiotensin-converting enzyme 2 (ACE2), through their inhibition of NLRP3 inflammasome activation and maximally reducing pyroptosis, indicating the basic principle behind the anti-inflammatory mechanism deployed by engineered MDEs [50].
MDEs or bacterial extracellular vesicles carry a diverse cargo, including proteins, lipids, nucleic acids such as mRNA, microRNA, and DNA, as well as microbe-associated molecular patterns (MAMPs). This cargo enables MDEs to modulate immune cell activity by interacting with pattern recognition receptors like TLR2, TLR4, and TLR9 on dendritic cells, macrophages, and B cells, often promoting tolerogenic responses from commensal sources while inducing pro-inflammatory cytokine release (e.g., TNF, IL-6, IL-1β) from pathogenic bacteria-derived MDEs. MDEs from probiotics like Bacteroides fragilis or Escherichia coli deliver polysaccharides or RNAs that enhance regulatory T cell differentiation and suppress excessive inflammation, supporting intestinal homeostasis and immunity. Thus, MDE cargo influences immune cell maturation, antigen presentation, and cytokine profiles, with probiotic MDEs shifting toward anti-inflammatory effects vital for mucosal tolerance [84,85,86].
MDEs act as carriers of miRNAs, which post-transcriptionally control host gene expression. Gut MDEs have the ability to travel to far-off organs, like the testes, where they change the expression of miRNA in circulating exosomes, which hinders spermatogenesis [66]. By activating or silencing particular mRNA targets, MDEs can affect physiological states through this cross-kingdom communication.
MDE cargo can promote the production of protective cytokines within cells. GELN-derived miRNAs target bacterial genes, upregulating metabolites that stimulate host production of Interleukin-22 (IL-22), a cytokine important for improving intestinal barrier function and treating colitis [62].

Figure 3: Mechanism of action for microbiota-derived exosomes (MDEs). (A) Entry: The vesicle enters the host cell via endocytosis or membrane fusion. (B) Intracellular action: Cargoes (miRNA/proteins) inhibit the NLRP3 inflammasome and NF-κB pathway. (C) Outcome: These interactions lead to gene regulation, reduced inflammation, and tissue repair. This original figure is prepared with https://BioRender.com.
6 Potential Therapeutic Applications of MDE
Microbiota-derived exosomes (including bacterial, gut microbiota, and probiotic EVs) are being explored mainly for immunomodulation, barrier repair, metabolic disease management, anti-tumor therapy, and as bioinspired drug delivery systems. Lactobacillus-derived exosomes show anti-inflammatory and immunomodulatory effects, improving experimental inflammatory bowel disease and other inflammatory conditions by modulating cytokines, barrier integrity, and gut microbiota [87]. MDEs can be used to treat polycystic ovary syndrome by reshaping gut microbiota composition, enhancing beneficial species, and suppressing pathobionts, thereby correcting metabolic-endocrine imbalance [88]. MDEs promote epithelial repair and tight-junction expression, supporting mucosal healing in IBD-like models and mitigating liver injury via gut–liver axis modulation. MDEs have been implicated in regulating the gut–brain axis and show promise in models of neurological disorders by modulating neuroinflammation and peripheral immune responses. They participate in microbiota–immune–brain signaling in neurodegenerative disease, suggesting use as both biomarkers and potential therapeutic modulators of microglial and peripheral immune activity [87]. MDEs can display antigens and immunostimulatory molecules on their surface, enabling their use as acellular vaccine platforms or adjuvants to enhance immune responses against pathogens [89]. Skin microbiota-derived EVs appear capable of modulating local immunity and barrier function, opening possibilities for topical immunotherapies or vaccines [90]. MDEs influence tumor immunity and responsiveness to immunotherapies by modulating systemic and tumor-associated immune landscapes, particularly along the gut–lung axis and in gastrointestinal and lung cancers [91]. Engineered MDEs can be loaded with chemotherapeutics (e.g., doxorubicin) or nucleic acids to achieve targeted delivery to tumors with low systemic toxicity [92].
7 Loading Therapeutics into MDEs
Intercellular communication and disease diagnosis depend on exosomes in major part. Drug delivery also benefits from these nanovesicles. Their therapeutic possibilities come from favorable characteristics, including low immunogenicity and great stability [52]. Their application is improved even more by a natural ability for cell-specific targeting. Studies are looking at exosomes from many sources for this purpose. Promising reservoirs for these platforms are both mesenchymal stem cells and plants [3,52]. They advocate the efficient treatment of infectious diseases and cancers, among other diseases. Furthermore, a major target for this technology is neurodegenerative diseases.
MDEs load therapeutic agents with a range of loading techniques. The chosen approach is much influenced by the cargo and intended usage. Presently, the most often used techniques are passive loading and electroporation. Cell pre-incubation is one new method that seems to enhance cargo integrity; the main success criteria in this regard are the type of drug and loading capacity. Commonly, incubated simply are hydrophobic drugs [93]. This passive method is based on diffusion across the exosomal membrane, but it frequently fails for hydrophilic compounds. The transient pores created by electroporation facilitate the entry of nucleic acids such as siRNA [94]. This strategy needs to be carefully optimized in order to prevent exosome aggregation. A good substitute is bovine lactoferrin molecular conjugates [95]. The effectiveness of electroporation for siRNA delivery is matched by these conjugates. Moreover, nanoparticle pre-incubation of cells shows higher loading efficiency than physicochemical techniques [96].
Exosomes are an extremely versatile platform for delivering both small molecules and proteins. In addition, exosomes can be utilized as delivery vehicles for RNA cargoes. Exosomes have several advantages compared to traditional methods of transporting biomaterials, such as their ability to cross biological barriers [97]. In addition, exosomes provide an innate transport mechanism for natural RNA and exhibit minimal immunogenicity. Plant exosomes have been shown to transmit small RNAs between plants and pathogens, with evidence that therapeutic small RNAs can be introduced into mammals via the use of plant exosome-like nanovesicles [98]. Thus, exosomes serve as a unique platform for RNA interference (RNAi) therapies [99,100]. Exosomes possess unique capabilities to deliver complex cargoes into specific tissues. In order for exosomes to be used in clinical applications, however, capacity limitations in their production must be addressed, and improvements in the targeting specificity of exosomes and efficiencies of RNA loading into exosomes must be made.
8 Engineering MDEs for Enhanced Drug Delivery
Functionalizing exosomes with an antibody ligand greatly enhances their therapeutic performance. This is true of Gut-derived MDEs. Peptides serve as a cost-effective alternative to antibodies, as well as being very reliable ligands. The use of phage display technology enables the selection of specific peptides that bind to a given target. This technology is being used extensively in the development of biomarker diagnostics for breast cancer [101]. Genetic engineering and covalent chemistry can both be used to change or modify the surfaces of MDEs. Non-covalent interactions, including aptamer-based modifications, can also contribute to the improved targeting of MDEs [102]. Unfortunately, targeting peptides used to functionalize MDEs can be at risk of undergoing proteolytic degradation during biogenesis. The addition of glycosylation motifs to targeting peptides can provide a significant increase in their successful delivery to neuroblastoma cells and increase their surface appearance [103]. Functionalization of MDEs with ligands provides an opportunity to more effectively deliver targeted therapeutic agents. Therefore, in order for peptide-based targeting to be successful, it is essential to develop methods to prevent peptide degradation. Continued bioengineering research is focused on optimizing the functionalization of exosomes [104].
Genetic engineering of source bacteria can drive hypervesiculation (e.g., peptidoglycan-modifying enzyme overexpression in Lactiplantibacillus plantarum increases BEV yield and supports oral IBD therapy in preclinical models) or enable surface display of targeting ligands (e.g., HER2-affibody fused to ClyA on Escherichia coli OMVs to deliver kinesin spindle protein (KSP)-siRNA selectively to HER2-positive cancer cells). Physicochemical loading methods include electroporation for doxorubicin or siRNA encapsulation, which can enhance tumor accumulation and antitumor efficacy compared with free drug or conventional carriers in CRC and other models, and surface chemistry (e.g., covalent ligand conjugation) to introduce additional targeting or functional moieties. Hybrid systems typically fuse OMVs with inorganic nanoparticles such as gold or with polymeric matrices (including hydrogels or mesoporous silica) to improve stability, tumor targeting, and controlled or stimuli-responsive release [105,106,107].
8.1 Genetic Engineering of Source Bacteria
Genetic modification of bacteria increases the production of extracellular vesicles. Lactiplantibacillus plantarum engineered to express a peptidoglycan-modifying enzyme produced bacterial extracellular vesicles with 66-fold higher yield. These vesicles reduced tissue damage in DSS-induced colitis more than wild-type L. plantarum cells. Escherichia coli outer membrane vesicles displaying HER2-affibody delivered KSP-siRNA and induced regression of HER2-positive tumors without toxicity. Bacterial extracellular vesicles enable CRISPR/Cas9 delivery for gene editing in neurodegeneration therapy [106,107,108].
8.2 Physicochemical and Surface Modifications
Chemotherapeutic drugs can be loaded into OMVs by electroporation, and doxorubicin-loaded OMVs have shown higher antitumor activity than liposomal doxorubicin in colorectal cancer models. Surface functionalization using click chemistry or aptamers can improve targeting and reduce nonspecific clearance, while glycosylation or other carbohydrate motifs can stabilize peptide cargos [105].
Gold nanoparticles can be engineered to cross the blood–brain barrier, and hybrid vesicle–nanoparticle platforms are under investigation to improve brain delivery. However, direct experimental evidence that outer membrane vesicle–gold nanoparticle hybrids enhance blood–brain barrier transport remains limited [109].
A variety of methods exist for engineering and attaching targeting ligands to the exosome surface. The first is to use custom-designed chemical modifications of the RNA nanoparticles in vitro and genetic modifications of the exosomes from cell lines. Chemical modifications of RNA nanoparticles allow the addition of tumor-targeting ligands to them. This also leads to a reduction in nonspecific binding via a mechanism involving electrostatic repulsion [110]. Another method has been to create hybrid nanocarriers formed by the fusion of liposomes and exosomes [111]. By creating hydrogels made from naturally occurring sources, these hydrogels can be combined with exosomes and provide controlled release of drugs [111]. The combination of exosomal membranes with synthetic nanoparticles (such as gold nanoparticles [AuNP]) may be a very effective way to fabricate targeted delivery of drugs or other therapeutics to specific tissues, including the brain [109,112]. The concept of “super homotypic targeting” will provide more efficient and homotypic targeting of cells through the use of exosomes. The presence of lanthanides on the surface of engineered exosomes can increase the effectiveness of cell-exosome interactions more than 25 times and permits cells of the same lineage to capture significantly more and faster exosomes than otherwise possible [113]. These various engineering strategies allow for the enhanced ability of MDEs to deliver drugs to a specific tissue. Therefore, the potential for engineered exosomes to assist in cancer diagnostics, immunotherapy, targeted drug delivery, and tissue engineering is significant.
Lipid-containing nanovesicles known as exosomes are becoming popular delivery vehicles. They are becoming important for gene editing technologies like clustered regularly interspaced short palindromic repeats (CRISPR)/Cas9. Long-distance transportation and biocompatibility are facilitated by these vesicles. They are good candidates for therapeutic editing because of their engineerability. It is possible to engineer exosomes to target particular cell types. They show how to package genome editing tools efficiently. This capability tackles issues in CRISPR/Cas9 clinical translation [114]. Experimental evidence indicates that extracellular vesicles can mediate horizontal transfer and stable integration of genomic DNA between eukaryotic cells in vitro. This raises the possibility that EV-mediated DNA transfer has contributed to genome evolution, although its in vivo and evolutionary relevance remains to be demonstrated [115]. This also poses the threat of potential off-target insertions during gene therapy. While the therapeutic potential of exosome-mediated gene-editing is exciting, it is important to be aware of possible risks and off-target effects that could be encountered when using this technology. In addition, the efficiency and specificity of exosome-based systems could provide a more attractive alternative to conventional viral vectors for gene therapy, which have shown limited cargo capacity and safety issues. Further exploration into exosome-based delivery systems, potentially in combination with carriers such as metal-organic frameworks (MOFs), may aid in the development of safe and efficient gene-editing technologies for clinical applications [116].
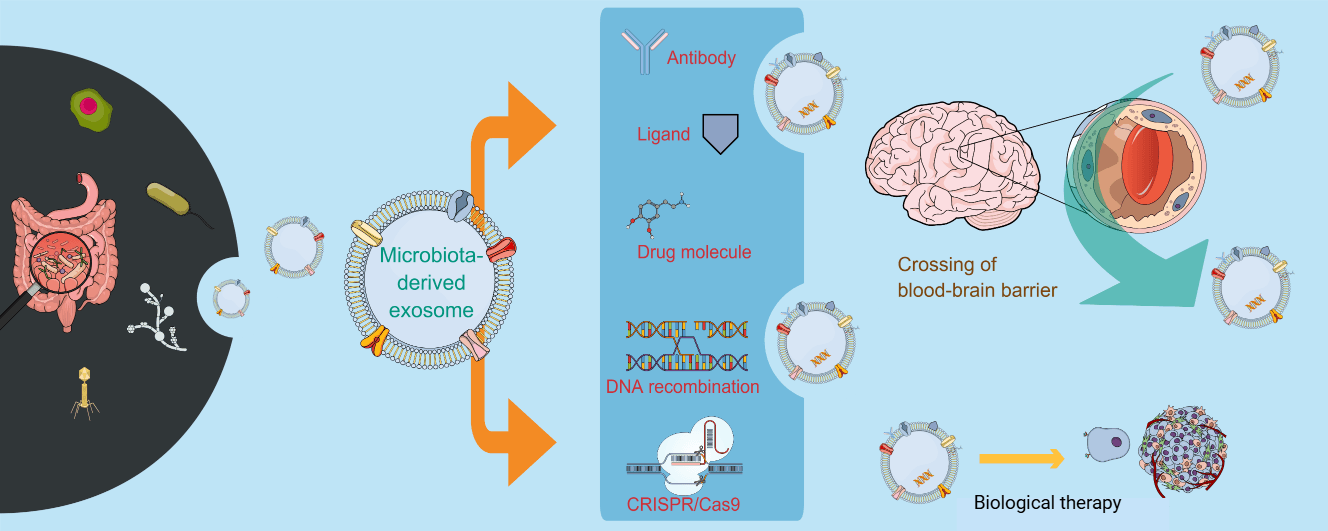
Engineered MDEs also show great promise in targeting genetic disorders and controlling gene expression. MDEs are naturally nontoxic, non-immunogenic, and biodegradable nanovesicles that are suitable for gene targeting. They can be decorated with particular surface ligands to achieve specific delivery to cancer or diseased tissues [117]. Indeed, exosomes derived from engineered HEK293T cells were able to target HER2-overexpressing breast cancer cells for therapy [118]. The crosstalk between microRNAs (miRNAs), intestinal microbiota, and intestinal immune system plays a critical role in maintaining intestinal homeostasis [119]. Endogenous and dietary miRNAs can impact microbial colonization and intestinal immunity, and thus represent a delivery cargo for engineered MDEs, which may be used to transfer specific miRNAs to control gene expression and homeostasis in the intestine [66,119]. These exosomes can carry a variety of biomolecules, including proteins, lipids, and nucleic acids, for gene regulation in target cells [67]. Furthermore, the recent development of genetic tools to study and manipulate non-model gut commensals, coupled with an ability to control these organisms, is now presenting new ways to modify the microbiome for the benefit of host physiology and treatment of disease [120]. Engineered exosomes are poised to play a significant role in personalized medicine and genetic disorder treatments. In summary, engineered MDEs offer a versatile platform for targeted gene therapy. Fig. 4 illustrates the engineering of MDEs for enhanced drug/gene delivery by surface functionalization and genetic engineering.

Figure 4: Engineering microbiota-derived exosomes (MDEs) for enhanced drug delivery. Strategies for modifying exosomes to improve therapeutic efficacy include exosome surface functionalization via attaching ligands by antibody, peptide, or protein binding and genetic engineering-based exosome expression of a protein or encapsulation of gene-editing tools, such as CRISPR/Cas9. This original figure is prepared with Mind the Graph (www.mindthegraph.com).
9.1 Scalability and Isolation Techniques
Current methods for isolating MDEs include ultracentrifugation, precipitation, filtration, and chromatography, each with benefits and scaling challenges [121]. Although ultracentrifugation is a commonly used technique, it is still time-consuming and not appropriate for large-scale production [122]. Helicobacter pylori MDEs have been isolated and characterized using a thorough methodology that has been documented. To separate intact OMVs, this purification procedure used size exclusion chromatography in conjunction with centrifugation and filtration. Western blotting and nanoparticle tracking analysis were used in subsequent characterization. Physical integrity was further validated by electron microscopy. The vesicles were enriched with bacterial components such as flagellin A (FlaA) and had an average size of 176.1 ± 4.7 nm [123].
A simple alternative that frequently compromises sample purity is polyethylene glycol (PEG) precipitation. At the same time, new microfluidic technologies allow for the efficient and quick isolation of exosomes. Isolating exosomes from small plasma volumes for point-of-care use is possible within 50 min using a double filtration microfluidic device [124]. Size exclusion and deterministic lateral displacement are two more microfluidic techniques. To isolate biologically active substances without using labels, methods utilizing viscoelastic and acoustic or inertial forces are employed [125]. Despite this, a major obstacle is the need to scale up production. Currently, therapeutic applications are hindered by exosome heterogeneity and low yield [126]. For exosomes to work better, better ways are needed to make them on a large scale and ways to load drugs into them [127]. Downstream analysis becomes more complicated when soluble components, such as cytokines, co-elute, making the purity of the sample essential [128]. As a result, manufacturing processes for MDEs must be standardized and scalable.
Exosome purification techniques have evolved greatly for MDEs such as those from Lactobacillus. However, a reliable technique is conventional differential ultracentrifugation. Recently added are density gradient centrifugation and size-exclusion chromatography. Moreover, practical methods based on membrane characteristics, including charge and hydrophobic interactions, exist. For exosomes produced from Lactobacillus, ultracentrifugation and size-exclusion chromatography have shown decent success [87,129]. Techniques of isolation can greatly affect the composition of exosome proteins and biological activity. A study on adipose-derived mesenchymal stem cells [130] revealed that different approaches affected protein profiles. This stresses the need to select appropriate methods for given goals. Standardized, reasonably priced purification methods define clinical application. Novel methods showing promise in increasing isolation efficiency are optimal ultrafiltration for urinary exosomes [131]. Examining the advantages of each method with respect to exosome characteristics is absolutely vital.
9.2 Purification Impacts on Immunomodulatory Cargo
MDE purification methods differentially preserve immunologically active cargos. Ultracentrifugation retains lipopolysaccharides and miRNAs critical for TLR signaling but risks protein denaturation. Size-exclusion chromatography maintains PSA/TLR2 ligands for Treg induction while removing inflammatory contaminants. Cargo loss during tangential flow filtration reduces NLRP3 inhibitory capacity by 40–60%, necessitating method optimization for clinical-grade MDEs. Purification strategies must balance yield, purity, and retention of immune-modulating lipids/proteins for therapeutic efficacy.
9.3 Safety and Regulatory Considerations
MDEs are pertinent players in host-microbe interactions, but their action bears consequences for the health of the host. Their disease-promoting capacity is of great concern. Gut microbiota exosomes transport bioactive components across physiological barriers [132]. Such boundaries that they are able to traverse are the intestinal and the blood-brain barrier. With respect to this transport capability, then, there is a further chance of bearing harmful encumbrances. Succinate increased gut bacterial lipopolysaccharides in extracellular vesicles in gastrointestinal tract-associated mastitis. These vesicles traversed across the damaged gut lining, invading the mammary glands. Inflammatory induction was via the TLR4/NF-κB signaling pathway [83], thereby underscoring MDEs as a double-edged sword: while they offer promising therapeutic avenues, they simultaneously possess the potential to induce harm. The growth of human hair was enhanced by EVs derived from exogenous microorganisms, including Lactobacillus species and stem cells [133]. These findings highlight the dual function of exosomes in the gut microbiota; hence, there is a necessity for increased safety assessments of any downstream therapeutics going forward. In this context, it is vital to better understand the mechanism/s of action on host cells in order to maximize clinical benefit.
Challenging translational barriers have delayed the clinical applicability of microbiota exosomes. There was no common protocol for isolation and characterization. Attainment of a gold standard is elusive [134]. Pertaining to the corrupting heterogeneity of exosomes, their products are not easily standardized and validated, a requirement for regulatory approval [135]. Regulatory hurdles are just as onerous as they may be for any exosome-based platform [134]. Compliance with good manufacturing practices (GMP) for clinicals makes for an additional obstacle [136]. Being aspectually varied, MDEs can raise the issue of how to clinically validate their functionality and efficacy in a safe manner across different disorders and further complicate regulatory acquisition [137]. Solutions to these challenges include establishing standardized protocols for isolation and characterization of cells, strictly adhering to GMP practices, and properly interpreting FDA guidance. Clearly defining clinical parameters will be critical for obtaining regulatory approval, and thereby progress cell therapy to the clinic [138].
10.1 Potential in Personalized and Precision Medicine
Interindividual differences of the microbiota are one of the important causes of the individual differences in drug efficacy and adverse reactions, so the treatment of people based on the same medication type would thus be more personalized [139,140]. On the basis of precision medicine, the use of the gut microbiota could effectively improve the therapeutic potential of numerous drugs and achieve a reduction in drug toxicity [139]. Furthermore, MDEs combined with the uniqueness of the exosomes and personal variation of physiological state may open a new field of drug delivery under personalization. This strategy could transform treatments for cancer, neurological disorders, and other diseases, improving efficacy and minimizing side effects [140,141]. Nonetheless, further research is required to address challenges in exosome isolation, purification, and targeted delivery for clinical applications.
MDEs are important mediators of intercellular communication. They have also been used to treat male infertility and dermatological conditions [142,143]. The gut microbiota has a profound impact on systemic health, and thus dysbiosis is implicated in the pathophysiology of a variety of disease states. There are challenges as well as opportunities for precision medicine, which arise because of the inter-individual variation in gut microbiota composition [141]. This has an impact on metabolic status, as well as how actively drugs targeting the microbiome are responded to by the body [140]. In dermatology, MDE-based treatments are durable, particularly in the context of alopecia. By actively promoting tissue regeneration and promoting follicles to form, these therapies will improve wound healing. In this regard, the fundamental principles of precision medicine are to be found in the tailoring of MDE therapy to specific patients. The capacity to analyze complex multi-omics data is provided by advances in computational technology, such as AI and machine learning. It is this kind of analytical capacity that permits the tailored development of therapies based on an individual patient’s microbiome. Nonetheless, there are still many problems to be resolved in terms of rigorous safety assessments and standardization processes. These approaches also need to be validated in long-term studies of efficacy [142]. The strategic application of MDEs to maximize their potential in personalized precision medicine is depicted in Fig. 5.

Figure 5: Illustration of exploring the power of microbiota-derived exosomes (MDEs) for personalized and precision medicine. (1) Microbiota profiling (2) Engineered MDE production (3) Targeted drug delivery (4) Personalized treatment outcome. This original figure is prepared with Mind the Graph (www.mindthegraph.com).
10.2 Opportunities for Innovation
New advancements in disciplines, including synthetic biology and nanotechnology, are changing the field of MDEs. These technologies improve applications in biomedicine significantly. Nanotechnology especially advances exosome-based drug delivery and diagnostics. The convergence of nanotechnology and synthetic biology has made the development of nano-artificial hybrid systems simpler [144]. This integration of synthetic and biological features maximizes biological performance. This type of cooperation makes it feasible to program RNA nanodevices inside living cells. These devices precisely control gene expression and accelerate the rates of chemical synthesis [145]. This multidisciplinary area is referred to as synthetic bionanotechnology. It effectively increases the possibilities of synthetic biology to improve the capacity of the microbiota exosomes. These advances are expected to bring new therapeutic strategies and better diagnostics. Nevertheless, ethical considerations and risk assessments remain critical.
The proteome of bacterial extracellular vesicles (bEVs) produced from the gut microbiome was investigated recently in a comparative case-control study. This work also looked at the makeup of the bacteria in whole faecal samples from healthy controls and solid tumours. The results revealed that bEVs were distinct taxonomic entities, unlike those of the parent gut microbiota. The bEV study also accurately categorized solid tumor patients. This performance exceeded the 93% accuracy rate among total faecal samples. These findings show how bEVs could be useful as cancer diagnostic biomarkers [146].
Recent advances in cancer immunotherapy show how crucial it is to combine MDEs with innovative treatment modalities. Exosomes obtained from immune cells offer a novel approach for changing the immune system to produce strong antitumor responses [48]. Natural exosomes do, however, have clinical limits in terms of their capacity to fight tumors and effectively deliver drugs. Thus, technological developments are needed to overcome these challenges. The gut flora greatly affects the modulation of the immune system. It affects the toxicity and potency of cancer treatments as well. The success of immunocheckpoint inhibitor treatment is linked to specific bacterial species [147]. Combining MDEs with immunotherapies thus has great potential to improve clinical outcomes. Combining gut microbiota modification with engineered exosomes could help to solve current constraints [48,49]. Novel methods looking at this synergy could simultaneously reduce side effects and raise effectiveness.
10.3 Areas Where Further Research Is Needed
MDE research is vital for drug delivery, particularly with regard to the need for production standardization. The large-scale production necessary for clinical use cannot be achieved with existing exosome isolation and purification methods [2,56]. Greater concentrations and purity require scalable, enhanced, optimized production [2]. There needs to be further characterization of the molecular composition and cargo of MDEs. Their biological properties and effects will be unlocked through a comprehensive characterization of their protein, mRNA, and microRNA contents. Further targeting of circulation time and efficiency is also needed. Therapeutic utility would be improved by bioengineering them to improve their targeting capacity and to extend their circulation lifetime [104]. Their drug delivery capacity could be enhanced by surface modification approaches. The efficacy and safety of drug delivery systems, including assessing immunogenicity, biocompatibility, and possible adverse effects if administered at high doses, are contingent upon in vivo research and clinical trials [2]. For oral drug delivery applications, research on their ability to traverse biological barriers, such as the intestinal epithelium, is also valuable.
MDEs can be described as ‘smart’ bio-inspired drug delivery technologies that exploit natural communication pathways (intercellular communication). MDEs possess a unique natural lipid composition for natural biocompatibility and the ability to navigate biological barriers by tight-junction stripping. These nanovesicles can predominantly be loaded with small molecules, RNA, and proteins as therapeutic agents that specifically target host cell receptors. MDEs modulate different important intracellular signaling pathways, including inflammatory and apoptotic pathways. MDEs can be engineered via surface functionalization or genetic alteration to further regulate the molecular interactions. However, standardization of production protocols and characterization of molecular cargoes must be ascertained for clinical translation. Engagement with MDEs as tools for personalized and precision medicine holds great potential for changing the way we combat cancer, inflammatory diseases, and male infertility, among others.
Acknowledgement:
Funding Statement: The Deanship of Scientific Research (DSR) at King Abdulaziz University (KAU), Jeddah, Saudi Arabia, has funded this project under grant No. (RG-20-166-43).
Author Contributions: Ibrahim M. Ibrahim: conceptualization, writing—original draft, writing—review & editing, funding acquisition. Shadab Md: conceptualization, supervision, resources, writing—original draft, writing—review & editing, funding acquisition. All authors reviewed and approved the final version of the manuscript.
Availability of Data and Materials: No data was used for the research described in the article.
Ethics Approval: Not applicable.
Conflicts of Interest: The authors declare no conflicts of interest.
References
1. Li S , Tang Y , Dou Y . The potential of milk-derived exosomes for drug delivery. Curr Drug Deliv. 2021; 18( 6): 688– 99. doi:10.2174/1567201817666200817112503. [Google Scholar] [CrossRef]
2. Cui Z , Amevor FK , Zhao X , Mou C , Pang J , Peng X , et al. Potential therapeutic effects of milk-derived exosomes on intestinal diseases. J Nanobiotechnol. 2023; 21( 1): 496. doi:10.1186/s12951-023-02176-8. [Google Scholar] [CrossRef]
3. Zhao W , Li K , Li L , Wang R , Lei Y , Yang H , et al. Mesenchymal stem cell-derived exosomes as drug delivery vehicles in disease therapy. Int J Mol Sci. 2024; 25( 14): 7715. doi:10.3390/ijms25147715. [Google Scholar] [CrossRef]
4. Hou X , Jiang H , Liu T , Yan J , Zhang F , Zhang X , et al. Depletion of gut microbiota resistance in 5×FAD mice enhances the therapeutic effect of mesenchymal stem cell-derived exosomes. Biomed Pharmacother. 2023; 161: 114455. doi:10.1016/j.biopha.2023.114455. [Google Scholar] [CrossRef]
5. Shen Y , Giardino Torchia ML , Lawson GW , Karp CL , Ashwell JD , Mazmanian SK . Outer membrane vesicles of a human commensal mediate immune regulation and disease protection. Cell Host Microbe. 2012; 12( 4): 509– 20. doi:10.1016/j.chom.2012.08.004. [Google Scholar] [CrossRef]
6. Yang J , Kim EK , McDowell A , Kim YK . Microbe-derived extracellular vesicles as a smart drug delivery system. Transl Clin Pharmacol. 2018; 26( 3): 103– 10. doi:10.12793/tcp.2018.26.3.103. [Google Scholar] [CrossRef]
7. Li Y , Zhou C , Liu H , Cai T , Fan H . Emerging roles of extracelluar vesicles derived from bacteria, mammalian or plant cells in the pathogenesis and clinical application of neurodegenerative diseases. Biomolecules. 2024; 14( 3): 312. doi:10.3390/biom14030312. [Google Scholar] [CrossRef]
8. Yang J , Shin TS , Kim JS , Jee YK , Kim YK . A new horizon of precision medicine: combination of the microbiome and extracellular vesicles. Exp Mol Med. 2022; 54( 4): 466– 82. doi:10.1038/s12276-022-00748-6. [Google Scholar] [CrossRef]
9. Nikfarjam S , Rezaie J , Zolbanin NM , Jafari R . Mesenchymal stem cell derived-exosomes: a modern approach in translational medicine. J Transl Med. 2020; 18( 1): 449. doi:10.1186/s12967-020-02622-3. [Google Scholar] [CrossRef]
10. Pandey SN , Afzal M , Goyal K , Rekha MM , Nayak PP , Maqbool M , et al. The role of MSC-derived exosomes in immune modulation and regenerative medicine. J Drug Target. 2026: 1– 19. doi:10.1080/1061186X.2026.2620707. [Google Scholar] [CrossRef]
11. Ha DH , Kim HK , Lee J , Kwon HH , Park GH , Yang SH , et al. Mesenchymal stem/stromal cell-derived exosomes for immunomodulatory therapeutics and skin regeneration. Cells. 2020; 9( 5): 1157. doi:10.3390/cells9051157. [Google Scholar] [CrossRef]
12. Tan Y , Luo X , Lv W , Hu W , Zhao C , Xiong M , et al. Tumor-derived exosomal components: the multifaceted roles and mechanisms in breast cancer metastasis. Cell Death Dis. 2021; 12( 6): 547. doi:10.1038/s41419-021-03825-2. [Google Scholar] [CrossRef]
13. Huang S , Dong M , Chen Q . Tumor-derived exosomes and their role in breast cancer metastasis. Int J Mol Sci. 2022; 23( 22): 13993. doi:10.3390/ijms232213993. [Google Scholar] [CrossRef]
14. Hazrati A , Soudi S , Malekpour K , Mahmoudi M , Rahimi A , Hashemi SM , et al. Immune cells-derived exosomes function as a double-edged sword: role in disease progression and their therapeutic applications. Biomark Res. 2022; 10( 1): 30. doi:10.1186/s40364-022-00374-4. [Google Scholar] [CrossRef]
15. Jin X , Zhang J , Zhang Y , He J , Wang M , Hei Y , et al. Different origin-derived exosomes and their clinical advantages in cancer therapy. Front Immunol. 2024; 15: 1401852. doi:10.3389/fimmu.2024.1401852. [Google Scholar] [CrossRef]
16. Huo L , Du X , Li X , Liu S , Xu Y . The emerging role of neural cell-derived exosomes in intercellular communication in health and neurodegenerative diseases. Front Neurosci. 2021; 15: 738442. doi:10.3389/fnins.2021.738442. [Google Scholar] [CrossRef]
17. Li F , Kang X , Xin W , Li X . The emerging role of extracellular vesicle derived from neurons/neurogliocytes in central nervous system diseases: novel insights into ischemic stroke. Front Pharmacol. 2022; 13: 890698. doi:10.3389/fphar.2022.890698. [Google Scholar] [CrossRef]
18. Prylińska-Jaśkowiak M , Kożuchowski M . The human gastroinstestinal tract microbiota in health–current knowledge summary. J Educ Health Sport. 2022; 12( 10): 41– 51. doi:10.12775/jehs.2022.12.10.005. [Google Scholar] [CrossRef]
19. Pevsner-Fischer M , Rot C , Tuganbaev T , Elinav E . The microbiota and its modulation in immune-mediated disorders. In: Immune rebalancing. Amsterdam, The Netherlands: Elsevier; 2016. p. 191– 227. doi:10.1016/b978-0-12-803302-9.00010-5. [Google Scholar] [CrossRef]
20. Wegierska AE , Charitos IA , Topi S , Potenza MA , Montagnani M , Santacroce L . The connection between physical exercise and gut microbiota: implications for competitive sports athletes. Sports Med. 2022; 52( 10): 2355– 69. doi:10.1007/s40279-022-01696-x. [Google Scholar] [CrossRef]
21. Jandhyala SM . Role of the normal gut microbiota. World J Gastroenterol. 2015; 21( 29): 8787. doi:10.3748/wjg.v21.i29.8787. [Google Scholar] [CrossRef]
22. Pham VT , Dold S , Rehman A , Bird JK , Steinert RE . Vitamins, the gut microbiome and gastrointestinal health in humans. Nutr Res. 2021; 95: 35– 53. doi:10.1016/j.nutres.2021.09.001. [Google Scholar] [CrossRef]
23. Xu X , Wang Z , Zhang X . The human microbiota associated with overall health. Crit Rev Biotechnol. 2015; 35( 1): 129– 40. doi:10.3109/07388551.2013.819485. [Google Scholar] [CrossRef]
24. Fan P , Bian B , Teng L , Nelson CD , Driver J , Elzo MA , et al. Host genetic effects upon the early gut microbiota in a bovine model with graduated spectrum of genetic variation. ISME J. 2020; 14( 1): 302– 17. doi:10.1038/s41396-019-0529-2. [Google Scholar] [CrossRef]
25. Krga I , Glibetic M . Gut microbiota in health and diseases. In: Comprehensive gut microbiota. Amsterdam, The Netherlands: Elsevier; 2022. p. 182– 98. doi:10.1016/b978-0-12-819265-8.00045-0. [Google Scholar] [CrossRef]
26. Duttaroy AK . Gut microbiota and the immune system and inflammation. In: Evidence-based nutrition and clinical evidence of bioactive foods in human health and disease. Amsterdam, The Netherlands: Elsevier; 2021. p. 311– 33. doi:10.1016/b978-0-12-822405-2.00031-1. [Google Scholar] [CrossRef]
27. Salzman NH . Microbiota–immune system interaction: an uneasy alliance. Curr Opin Microbiol. 2011; 14( 1): 99– 105. doi:10.1016/j.mib.2010.09.018. [Google Scholar] [CrossRef]
28. Shi N , Li N , Duan X , Niu H . Interaction between the gut microbiome and mucosal immune system. Mil Med Res. 2017; 4: 14. doi:10.1186/s40779-017-0122-9. [Google Scholar] [CrossRef]
29. Cañas MA , Fábrega MJ , Giménez R , Badia J , Baldomà L . Outer membrane vesicles from probiotic and commensal Escherichia coli activate NOD1-mediated immune responses in intestinal epithelial cells. Front Microbiol. 2018; 9: 498. doi:10.3389/fmicb.2018.00498. [Google Scholar] [CrossRef]
30. Saini A , Dalal P , Sharma D . Deciphering the interdependent labyrinth between gut microbiota and the immune system. Lett Appl Microbiol. 2022; 75( 5): 1122– 35. doi:10.1111/lam.13775. [Google Scholar] [CrossRef]
31. Wang Z , Gong J . Gut microbiota and immune-related diseases. Chin J Gastrointest Surg. 2022; 25: 777– 83. doi:10.3760/cma.j.cn441530-20211130-00480. [Google Scholar] [CrossRef]
32. Ma PJ , Wang MM , Wang Y . Gut microbiota: a new insight into lung diseases. Biomed Pharmacother. 2022; 155: 113810. doi:10.1016/j.biopha.2022.113810. [Google Scholar] [CrossRef]
33. Cuesta CM , Guerri C , Ureña J , Pascual M . Role of microbiota-derived extracellular vesicles in gut-brain communication. Int J Mol Sci. 2021; 22( 8): 4235. doi:10.3390/ijms22084235. [Google Scholar] [CrossRef]
34. Peng Y , Yin S , Wang M . Extracellular vesicles of bacteria as potential targets for immune interventions. Hum Vaccin Immunother. 2021; 17( 3): 897– 903. doi:10.1080/21645515.2020.1799667. [Google Scholar] [CrossRef]
35. Liu BD , Akbar R , Oliverio A , Thapa K , Wang X , Fan GC . Bacterial extracellular vesicles in the regulation of inflammatory response and host-microbe interactions. Shock. 2024; 61( 2): 175– 88. doi:10.1097/SHK.0000000000002252. [Google Scholar] [CrossRef]
36. Melo-Marques I , Cardoso SM , Empadinhas N . Bacterial extracellular vesicles at the interface of gut microbiota and immunity. Gut Microbes. 2024; 16( 1): 2396494. doi:10.1080/19490976.2024.2396494. [Google Scholar] [CrossRef]
37. Fonseca S , Carvalho AL , Miquel-Clopés A , Jones EJ , Juodeikis R , Stentz R , et al. Extracellular vesicles produced by the human gut commensal bacterium Bacteroides thetaiotaomicron elicit anti-inflammatory responses from innate immune cells. Front Microbiol. 2022; 13: 1050271. doi:10.3389/fmicb.2022.1050271. [Google Scholar] [CrossRef]
38. Groot Kormelink T , Mol S , de Jong EC , Wauben MHM . The role of extracellular vesicles when innate meets adaptive. Semin Immunopathol. 2018; 40( 5): 439– 52. doi:10.1007/s00281-018-0681-1. [Google Scholar] [CrossRef]
39. Wu Q , Kan J , Fu C , Liu X , Cui Z , Wang S , et al. Insights into the unique roles of extracellular vesicles for gut health modulation: mechanisms, challenges, and perspectives. Curr Res Microb Sci. 2024; 7: 100301. doi:10.1016/j.crmicr.2024.100301. [Google Scholar] [CrossRef]
40. Alberti G , Mazzola M , Gagliardo C , Pitruzzella A , Fucarini A , Giammanco M , et al. Extracellular vesicles derived from gut microbiota in inflammatory bowel disease and colorectal cancer. Biomed Pap Med Fac Univ Palacky Olomouc Czech Repub. 2021; 165( 3): 233– 40. doi:10.5507/bp.2021.042. [Google Scholar] [CrossRef]
41. Haas-Neill S , Forsythe P . A budding relationship: bacterial extracellular vesicles in the microbiota-gut-brain axis. Int J Mol Sci. 2020; 21( 23): 8899. doi:10.3390/ijms21238899. [Google Scholar] [CrossRef]
42. Du X , Ley R , Buck AH . microRNAs and extracellular vesicles in the gut: new host modulators of the microbiome? Microlife. 2021; 2: uqab010. doi:10.1093/femsml/uqab010. [Google Scholar] [CrossRef]
43. Comito D , Cascio A , Romano C . Microbiota biodiversity in inflammatory bowel disease. Ital J Pediatr. 2014; 40: 32. doi:10.1186/1824-7288-40-32. [Google Scholar] [CrossRef]
44. Quaglio AEV , Grillo TG , De Oliveira ECS , Di Stasi LC , Sassaki LY . Gut microbiota, inflammatory bowel disease and colorectal cancer. World J Gastroenterol. 2022; 28( 30): 4053– 60. doi:10.3748/wjg.v28.i30.4053. [Google Scholar] [CrossRef]
45. Foppa C , Rizkala T , Repici A , Hassan C , Spinelli A . Microbiota and IBD: current knowledge and future perspectives. Dig Liver Dis. 2024; 56( 6): 911– 22. doi:10.1016/j.dld.2023.11.015. [Google Scholar] [CrossRef]
46. Zhang M , Sun K , Wu Y , Yang Y , Tso P , Wu Z . Interactions between intestinal microbiota and host immune response in inflammatory bowel disease. Front Immunol. 2017; 8: 942. doi:10.3389/fimmu.2017.00942. [Google Scholar] [CrossRef]
47. Poletti M , Arnauts K , Ferrante M , Korcsmaros T . Organoid-based models to study the role of host-microbiota interactions in IBD. J Crohn’s Colitis. 2021; 15( 7): 1222– 35. doi:10.1093/ecco-jcc/jjaa257. [Google Scholar] [CrossRef]
48. Jung I , Shin S , Baek MC , Yea K . Modification of immune cell-derived exosomes for enhanced cancer immunotherapy: current advances and therapeutic applications. Exp Mol Med. 2024; 56( 1): 19– 31. doi:10.1038/s12276-023-01132-8. [Google Scholar] [CrossRef]
49. Soto Chervin C , Gajewski TF . Microbiome-based interventions: therapeutic strategies in cancer immunotherapy. Immunooncol Technol. 2020; 8: 12– 20. doi:10.1016/j.iotech.2020.11.001. [Google Scholar] [CrossRef]
50. Yang S , Cao J , Wang Y , Chen Q , Li F , Gao Y , et al. Small intestinal endocrine cell derived exosomal ACE2 protects islet β-cell function by inhibiting the activation of NLRP3 inflammasome and reducing β-cell pyroptosis. Int J Nanomedicine. 2024; 19: 4957– 76. doi:10.2147/IJN.S450337. [Google Scholar] [CrossRef]
51. Ahuja A , Dhanalekshmi UM , Saraswathy MP , Nandakumar S , Arul Prakash F , Gurpreet KN . Role of the gut microbiome in diabetes and cardiovascular DiseasesIncluding restoration and targeting approaches—a review. Drug Metab Bioanal Lett. 2022; 15( 3): 133– 49. doi:10.2174/2949681015666220615120300. [Google Scholar] [CrossRef]
52. Batrakova EV , Kim MS . Using exosomes, naturally-equipped nanocarriers, for drug delivery. J Control Release. 2015; 219: 396– 405. doi:10.1016/j.jconrel.2015.07.030. [Google Scholar] [CrossRef]
53. Rao D , Huang D , Sang C , Zhong T , Zhang Z , Tang Z . Advances in mesenchymal stem cell-derived exosomes as drug delivery vehicles. Front Bioeng Biotechnol. 2022; 9: 797359. doi:10.3389/fbioe.2021.797359. [Google Scholar] [CrossRef]
54. Manandhar S , Kothandan VK , Oh J , Yoo SH , Hwang J , Hwang SR . A pharmaceutical investigation into exosomes. J Pharm Investig. 2018; 48( 6): 617– 26. doi:10.1007/s40005-018-0391-7. [Google Scholar] [CrossRef]
55. Madhan S , Dhar R , Devi A . Plant-derived exosomes: a green approach for cancer drug delivery. J Mater Chem B. 2024; 12( 9): 2236– 52. doi:10.1039/d3tb02752j. [Google Scholar] [CrossRef]
56. Rajput A , Varshney A , Bajaj R , Pokharkar V . Exosomes as new generation vehicles for drug delivery: biomedical applications and future perspectives. Molecules. 2022; 27( 21): 7289. doi:10.3390/molecules27217289. [Google Scholar] [CrossRef]
57. Zhao Y , Liu T , Zhou M . Immune-cell-derived exosomes for cancer therapy. Mol Pharmaceutics. 2022; 19( 9): 3042– 56. doi:10.1021/acs.molpharmaceut.2c00407. [Google Scholar] [CrossRef]
58. Spiljar M , Merkler D , Trajkovski M . The immune system bridges the gut microbiota with systemic energy homeostasis: focus on TLRs, mucosal barrier, and SCFAs. Front Immunol. 2017; 8: 1353. doi:10.3389/fimmu.2017.01353. [Google Scholar] [CrossRef]
59. Zou Z , Li H , Xu G , Hu Y , Zhang W , Tian K . Current knowledge and future perspectives of exosomes as nanocarriers in diagnosis and treatment of diseases. Int J Nanomed. 2023; 18: 4751– 78. doi:10.2147/IJN.S417422. [Google Scholar] [CrossRef]
60. Li C , Qin S , Wen Y , Zhao W , Huang Y , Liu J . Overcoming the blood-brain barrier: exosomes as theranostic nanocarriers for precision neuroimaging. J Control Release. 2022; 349: 902– 16. doi:10.1016/j.jconrel.2022.08.002. [Google Scholar] [CrossRef]
61. Zhang M , Zang X , Wang M , Li Z , Qiao M , Hu H , et al. Exosome-based nanocarriers as bio-inspired and versatile vehicles for drug delivery: recent advances and challenges. J Mater Chem B. 2019; 7( 15): 2421– 33. doi:10.1039/c9tb00170k. [Google Scholar] [CrossRef]
62. Teng Y , Ren Y , Sayed M , Hu X , Lei C , Kumar A , et al. Plant-derived exosomal microRNAs shape the gut microbiota. Cell Host Microbe. 2018; 24( 5): 637– 52.e8. doi:10.1016/j.chom.2018.10.001. [Google Scholar] [CrossRef]
63. Le HH , Lee MT , Besler KR , Johnson EL . Host hepatic metabolism is modulated by gut microbiota-derived sphingolipids. Cell Host Microbe. 2022; 30( 6): 798– 808.e7. doi:10.1016/j.chom.2022.05.002. [Google Scholar] [CrossRef]
64. Kandalgaonkar MR , Kumar V , Vijay-Kumar M . Digestive dynamics: unveiling interplay between the gut microbiota and the liver in macronutrient metabolism and hepatic metabolic health. Physiol Rep. 2024; 12( 12): e16114. doi:10.14814/phy2.16114. [Google Scholar] [CrossRef]
65. Dong X , Liu Y , Yang X , Li T . Extracellular vesicle miRNAs as key mediators in diet-gut microbiome-host interplay. Trends Food Sci Technol. 2023; 136: 268– 81. doi:10.1016/j.tifs.2023.05.005. [Google Scholar] [CrossRef]
66. Chen T , Zhang B , He G , Wang N , Cao M , Shen C , et al. Gut-derived exosomes mediate the microbiota dysbiosis-induced spermatogenesis impairment by targeting meioc in mice. Adv Sci. 2024; 11( 22): 2310110. doi:10.1002/advs.202310110. [Google Scholar] [CrossRef]
67. Kim H , Kim EH , Kwak G , Chi SG , Kim SH , Yang Y . Exosomes: cell-derived nanoplatforms for the delivery of cancer therapeutics. Int J Mol Sci. 2020; 22( 1): 14. doi:10.3390/ijms22010014. [Google Scholar] [CrossRef]
68. Lin Y , Lu Y , Li X . Biological characteristics of exosomes and genetically engineered exosomes for the targeted delivery of therapeutic agents. J Drug Target. 2020; 28( 2): 129– 41. doi:10.1080/1061186X.2019.1641508. [Google Scholar] [CrossRef]
69. Vashisht M , Rani P , Onteru SK , Singh D . Curcumin encapsulated in milk exosomes resists human digestion and possesses enhanced intestinal permeability in vitro. Appl Biochem Biotechnol. 2017; 183( 3): 993– 1007. doi:10.1007/s12010-017-2478-4. [Google Scholar] [CrossRef]
70. Das C , Jena BC , Banerjee I , Das S , Parekh A , Bhutia SK , et al. Exosome as a novel shuttle for delivery of therapeutics across biological barriers. Mol Pharmaceutics. 2019; 16( 1): 24– 40. doi:10.1021/acs.molpharmaceut.8b00901. [Google Scholar] [CrossRef]
71. Osaid Z , Haider M , Hamoudi R , Harati R . Exosomes interactions with the blood-brain barrier: implications for cerebral disorders and therapeutics. Int J Mol Sci. 2023; 24( 21): 15635. doi:10.3390/ijms242115635. [Google Scholar] [CrossRef]
72. Elliott RO , He M . Unlocking the power of exosomes for crossing biological barriers in drug delivery. Pharmaceutics. 2021; 13( 1): 122. doi:10.3390/pharmaceutics13010122. [Google Scholar] [CrossRef]
73. Liu S , Gao J , Liu K , Zhang HL . Microbiota-gut-brain axis and Alzheimer’s disease: implications of the blood-brain barrier as an intervention target. Mech Ageing Dev. 2021; 199: 111560. doi:10.1016/j.mad.2021.111560. [Google Scholar] [CrossRef]
74. Knox EG , Aburto MR , Tessier C , Nagpal J , Clarke G , O’Driscoll CM , et al. Microbial-derived metabolites induce actin cytoskeletal rearrangement and protect blood-brain barrier function. iScience. 2022; 25( 12): 105648. doi:10.1016/j.isci.2022.105648. [Google Scholar] [CrossRef]
75. Al-Asmakh M , Hedin L . Microbiota and the control of blood-tissue barriers. Tissue Barriers. 2015; 3( 3): e1039691. doi:10.1080/21688370.2015.1039691. [Google Scholar] [CrossRef]
76. Pan J , Wang Z , Huang X , Xue J , Zhang S , Guo X , et al. Bacteria-derived outer-membrane vesicles hitchhike neutrophils to enhance ischemic stroke therapy. Adv Mater. 2023; 35( 38): e2301779. doi:10.1002/adma.202301779. [Google Scholar] [CrossRef]
77. Palacios E , Lobos-González L , Guerrero S , Kogan MJ , Shao B , Heinecke JW , et al. Helicobacter pylori outer membrane vesicles induce astrocyte reactivity through nuclear factor-κappa B activation and cause neuronal damage in vivo in a murine model. J Neuroinflammation. 2023; 20( 1): 66. doi:10.1186/s12974-023-02728-7. [Google Scholar] [CrossRef]
78. Fang Y , Wang Z , Liu X , Tyler BM . Biogenesis and biological functions of extracellular vesicles in cellular and organismal communication with microbes. Front Microbiol. 2022; 13: 817844. doi:10.3389/fmicb.2022.817844. [Google Scholar] [CrossRef]
79. Ribovski L , Joshi B , Gao J , Zuhorn I . Breaking free: endocytosis and endosomal escape of extracellular vesicles. Extracell Vesicles Circ Nucl Acids. 2023; 4( 2): 283– 305. doi:10.20517/evcna.2023.26. [Google Scholar] [CrossRef]
80. Gnopo YMD , Misra A , Hsu HL , DeLisa MP , Daniel S , Putnam D . Induced fusion and aggregation of bacterial outer membrane vesicles: experimental and theoretical analysis. J Colloid Interface Sci. 2020; 578: 522– 32. doi:10.1016/j.jcis.2020.04.068. [Google Scholar] [CrossRef]
81. Hays KE , Pfaffinger JM , Ryznar R . The interplay between gut microbiota, short-chain fatty acids, and implications for host health and disease. Gut Microbes. 2024; 16( 1): 2393270. doi:10.1080/19490976.2024.2393270. [Google Scholar] [CrossRef]
82. Liu SH , Yang XF , Liang L , Song BB , Song XM , Yang YJ , et al. Regulatory mechanisms of the gut microbiota-short chain fatty acids signaling axis in slow transit constipation and progress in multi-target interventions. Front Microbiol. 2025; 16: 1689597. doi:10.3389/fmicb.2025.1689597. [Google Scholar] [CrossRef]
83. Qiu M , Zhang N , Zou C , Xie J , Zhao C , Tang R , et al. Succinate exacerbates mastitis in mice via gut microbiota-derived extracellular vesicles. ResearchSquare. 2023. doi:10.21203/rs.3.rs-3585412/v1. [Google Scholar] [CrossRef]
84. Martínez-Ruiz S , Sáez-Fuertes L , Casanova-Crespo S , Rodríguez-Lagunas MJ , Pérez-Cano FJ , Badia J , et al. Microbiota-derived extracellular vesicles promote immunity and intestinal maturation in suckling rats. Nutrients. 2023; 15( 21): 4701. doi:10.3390/nu15214701. [Google Scholar] [CrossRef]
85. Hassan A , Huang Y , Mahjabina , Sudiro OH , Wang X , He J , et al. Emerging role of gut microbiota extracellular vesicle in neurodegenerative disorders and insights on their therapeutic management. iMetaOmics. 2024; 1( 2): e33. doi:10.1002/imo2.33. [Google Scholar] [CrossRef]
86. Macia L , Nanan R , Hosseini-Beheshti E , Grau GE . Host- and microbiota-derived extracellular vesicles, immune function, and disease development. Int J Mol Sci. 2019; 21( 1): 107. doi:10.3390/ijms21010107. [Google Scholar] [CrossRef]
87. Liu R . A promising area of research in medicine: recent advances in properties and applications of Lactobacillus-derived exosomes. Front Microbiol. 2024; 15: 1266510. doi:10.3389/fmicb.2024.1266510. [Google Scholar] [CrossRef]
88. Yang L , Liu T , Liao Y , Ren Y , Zheng Z , Zhang M , et al. Potential therapeutic application and mechanism of gut microbiota-derived extracellular vesicles in polycystic ovary syndrome. Biomed Pharmacother. 2024; 180: 117504. doi:10.1016/j.biopha.2024.117504. [Google Scholar] [CrossRef]
89. Liu H , Zhang H , Han Y , Hu Y , Geng Z , Su J . Bacterial extracellular vesicles-based therapeutic strategies for bone and soft tissue tumors therapy. Theranostics. 2022; 12( 15): 6576– 94. doi:10.7150/thno.78034. [Google Scholar] [CrossRef]
90. Rajan TS , Saiganesh R , Sivagnanavelmurugan M , Diomede F . Human skin microbiota-derived extracellular vesicles and their cosmeceutical possibilities—a mini review. Exp Dermatol. 2025; 34( 3): e70073. doi:10.1111/exd.70073. [Google Scholar] [CrossRef]
91. Albano GD , Taverna S . From microbiota to cancer: role of extracellular vesicles in gut-lung axis. Cancers. 2025; 17( 24): 3946. doi:10.3390/cancers17243946. [Google Scholar] [CrossRef]
92. Kumar MA , Baba SK , Sadida HQ , Al Marzooqi S , Jerobin J , Altemani FH , et al. Extracellular vesicles as tools and targets in therapy for diseases. Signal Transduct Target Ther. 2024; 9( 1): 27. doi:10.1038/s41392-024-01735-1. [Google Scholar] [CrossRef]
93. Xi XM , Xia SJ , Lu R . Drug loading techniques for exosome-based drug delivery systems. Pharmazie. 2021; 76( 2): 61– 7. doi:10.1691/ph.2021.0128. [Google Scholar] [CrossRef]
94. Familtseva A , Jeremic N , Tyagi SC . Exosomes: cell-created drug delivery systems. Mol Cell Biochem. 2019; 459( 1): 1– 6. doi:10.1007/s11010-019-03545-4. [Google Scholar] [CrossRef]
95. Shandilya S , Rani P , Onteru SK , Singh D . Natural ligand-receptor mediated loading of siRNA in milk derived exosomes. J Biotechnol. 2020; 318: 1– 9. doi:10.1016/j.jbiotec.2020.04.015. [Google Scholar] [CrossRef]
96. Sancho-Albero M , Del Mar Encabo-Berzosa M , Beltrán-Visiedo M , Fernández-Messina L , Sebastián V , Sánchez-Madrid F , et al. Efficient encapsulation of theranostic nanoparticles in cell-derived exosomes: leveraging the exosomal biogenesis pathway to obtain hollow gold nanoparticle-hybrids. Nanoscale. 2019; 11( 40): 18825– 36. doi:10.1039/c9nr06183e. [Google Scholar] [CrossRef]
97. Shahabipour F , Barati N , Johnston TP , Derosa G , Maffioli P , Sahebkar A . Exosomes: nanoparticulate tools for RNA interference and drug delivery. J Cell Physiol. 2017; 232( 7): 1660– 8. doi:10.1002/jcp.25766. [Google Scholar] [CrossRef]
98. Chen YX , Cai Q . Plant exosome-like nanovesicles and their role in the innovative delivery of RNA therapeutics. Biomedicines. 2023; 11( 7): 1806. doi:10.3390/biomedicines11071806. [Google Scholar] [CrossRef]
99. Zhou Y , Zhou G , Tian C , Jiang W , Jin L , Zhang C , et al. Exosome-mediated small RNA delivery for gene therapy. Wires RNA. 2016; 7( 6): 758– 71. doi:10.1002/wrna.1363. [Google Scholar] [CrossRef]
100. Duechler M . Vehicles for small interfering RNA transfection: exosomes versus synthetic nanocarriers. DNA RNA Nanotechnol. 2013; 1( 1): 16– 26. doi:10.2478/rnan-2013-0002. [Google Scholar] [CrossRef]
101. da Fonseca Alves R , Pallarès-Rusiñol A , Rossi R , Martí M , Vaz ER , de Araújo TG , et al. Peptide-based biosensing approaches for targeting breast cancer-derived exosomes. Biosens Bioelectron. 2024; 255: 116211. doi:10.1016/j.bios.2024.116211. [Google Scholar] [CrossRef]
102. Salunkhe S , Dheeraj , Basak M , Chitkara D , Mittal A . Surface functionalization of exosomes for target-specific delivery and in vivo imaging & tracking: strategies and significance. J Control Release. 2020; 326: 599– 614. doi:10.1016/j.jconrel.2020.07.042. [Google Scholar] [CrossRef]
103. Hung ME , Leonard JN . Stabilization of exosome-targeting peptides via engineered glycosylation. J Biol Chem. 2015; 290( 13): 8166– 72. doi:10.1074/jbc.M114.621383. [Google Scholar] [CrossRef]
104. Sadeghi S , Tehrani FR , Tahmasebi S , Shafiee A , Hashemi SM . Exosome engineering in cell therapy and drug delivery. Inflammopharmacology. 2023; 31( 1): 145– 69. doi:10.1007/s10787-022-01115-7. [Google Scholar] [CrossRef]
105. Zheng K , Feng Y , Li L , Kong F , Gao J , Kong X . Engineered bacterial outer membrane vesicles: a versatile bacteria-based weapon against gastrointestinal tumors. Theranostics. 2024; 14( 2): 761– 87. doi:10.7150/thno.85917. [Google Scholar] [CrossRef]
106. Gujrati V , Kim S , Kim SH , Min JJ , Choy HE , Kim SC , et al. Bioengineered bacterial outer membrane vesicles as cell-specific drug-delivery vehicles for cancer therapy. ACS Nano. 2014; 8( 2): 1525– 37. doi:10.1021/nn405724x. [Google Scholar] [CrossRef]
107. Pirolli NH , Levy D , Schledwitz A , Moura NS , Solomon TJ , Powsner EH , et al. Genetically-programmed hypervesiculation of lactiplantibacillus plantarum increases production of bacterial extracellular vesicles with therapeutic efficacy in a preclinical inflammatory bowel disease model. bioRxiv. 2025. doi:10.1101/2025.06.20.660770. [Google Scholar] [CrossRef]
108. Srivastava P , Kim KS . Membrane vesicles derived from gut microbiota and probiotics: cutting-edge therapeutic approaches for multidrug-resistant superbugs linked to neurological anomalies. Pharmaceutics. 2022; 14( 11): 2370. doi:10.3390/pharmaceutics14112370. [Google Scholar] [CrossRef]
109. Khongkow M , Yata T , Boonrungsiman S , Ruktanonchai UR , Graham D , Namdee K . Surface modification of gold nanoparticles with neuron-targeted exosome for enhanced blood-brain barrier penetration. Sci Rep. 2019; 9( 1): 8278. doi:10.1038/s41598-019-44569-6. [Google Scholar] [CrossRef]
110. Uddin N , Binzel DW , Shu D , Fu TM , Guo P . Targeted delivery of RNAi to cancer cells using RNA-ligand displaying exosome. Acta Pharm Sin B. 2023; 13( 4): 1383– 99. doi:10.1016/j.apsb.2022.11.019. [Google Scholar] [CrossRef]
111. Elkhoury K , Koçak P , Kang A , Arab-Tehrany E , Ellis Ward J , Shin SR . Engineering smart targeting nanovesicles and their combination with hydrogels for controlled drug delivery. Pharmaceutics. 2020; 12( 9): 849. doi:10.3390/pharmaceutics12090849. [Google Scholar] [CrossRef]
112. Soltanmohammadi F , Gharehbaba AM , Zangi AR , Adibkia K , Javadzadeh Y . Current knowledge of hybrid nanoplatforms composed of exosomes and organic/inorganic nanoparticles for disease treatment and cell/tissue imaging. Biomed Pharmacother. 2024; 178: 117248. doi:10.1016/j.biopha.2024.117248. [Google Scholar] [CrossRef]
113. Wang HS , Umirbaeva A , Zhou Y , Yang ZW , Hayashi M , Ishii N , et al. Super homotypic targeting by exosome surface engineering. bioRxiv. 2024. doi:10.1101/2024.05.01.592036. [Google Scholar] [CrossRef]
114. Duan L , Ouyang K , Wang J , Xu L , Xu X , Wen C , et al. Exosomes as targeted delivery platform of CRISPR/Cas9 for therapeutic genome editing. ChemBioChem. 2021; 22( 24): 3360– 8. doi:10.1002/cbic.202100359. [Google Scholar] [CrossRef]
115. Fischer S , Cornils K , Speiseder T , Badbaran A , Reimer R , Indenbirken D , et al. Indication of horizontal DNA gene transfer by extracellular vesicles. PLoS One. 2016; 11( 9): e0163665. doi:10.1371/journal.pone.0163665. [Google Scholar] [CrossRef]
116. Shaikhutdinov IH , Ilyasov PV , Gribkova OV , Limareva LV . Non-viral systems for intracellular delivery of genome editing tools. Vavilovskii Zhurnal Genet Selektsii. 2024; 28( 2): 239– 48. doi:10.18699/vjgb-24-28. [Google Scholar] [CrossRef]
117. Chronopoulos A , Kalluri R . Emerging role of bacterial extracellular vesicles in cancer. Oncogene. 2020; 39( 46): 6951– 60. doi:10.1038/s41388-020-01509-3. [Google Scholar] [CrossRef]
118. Limoni SK , Moghadam MF , Moazzeni SM , Gomari H , Salimi F . Engineered exosomes for targeted transfer of siRNA to HER2 positive breast cancer cells. Appl Biochem Biotechnol. 2019; 187( 1): 352– 64. doi:10.1007/s12010-018-2813-4. [Google Scholar] [CrossRef]
119. Bi K , Zhang X , Chen W , Diao H . microRNAs regulate intestinal immunity and gut microbiota for gastrointestinal health: a comprehensive review. Genes. 2020; 11( 9): 1075. doi:10.3390/genes11091075. [Google Scholar] [CrossRef]
120. Jin WB , Li TT , Huo D , Qu S , Li XV , Arifuzzaman M , et al. Genetic manipulation of gut microbes enables single-gene interrogation in a complex microbiome. Cell. 2022; 185( 3): 547– 62.e22. doi:10.1016/j.cell.2021.12.035. [Google Scholar] [CrossRef]
121. Shtam T , Evtushenko V , Samsonov R , Zabrodskaya Y , Kamyshinsky R , Zabegina L , et al. Evaluation of immune and chemical precipitation methods for plasma exosome isolation. PLoS One. 2020; 15( 11): e0242732. doi:10.1371/journal.pone.0242732. [Google Scholar] [CrossRef]
122. Skottvoll FS , Berg HE , Bjørseth K , Lund K , Roos N , Bekhradnia S , et al. Ultracentrifugation versus kit exosome isolation: nanoLC-MS and other tools reveal similar performance biomarkers, but also contaminations. Future Sci OA. 2018; 5( 1): FSO359. doi:10.4155/fsoa-2018-0088. [Google Scholar] [CrossRef]
123. Xie J , Cools L , Van Imschoot G , Van Wonterghem E , Pauwels MJ , Vlaeminck I , et al. Helicobacter pylori-derived outer membrane vesicles contribute to Alzheimer’s disease pathogenesis via C3-C3aR signalling. J Extracell Vesicles. 2023; 12( 2): 12306. doi:10.1002/jev2.12306. [Google Scholar] [CrossRef]
124. Ramnauth N , Neubarth E , Makler-Disatham A , Sher M , Soini S , Merk V , et al. Development of a microfluidic device for exosome isolation in point-of-care settings. Sensors. 2023; 23( 19): 8292. doi:10.3390/s23198292. [Google Scholar] [CrossRef]
125. Hassanpour Tamrin S , Sanati Nezhad A , Sen A . Label-free isolation of exosomes using microfluidic technologies. ACS Nano. 2021; 15( 11): 17047– 79. doi:10.1021/acsnano.1c03469. [Google Scholar] [CrossRef]
126. Yamashita T , Takahashi Y , Takakura Y . Possibility of exosome-based therapeutics and challenges in production of exosomes eligible for therapeutic application. Biol Pharm Bull. 2018; 41( 6): 835– 42. doi:10.1248/bpb.b18-00133. [Google Scholar] [CrossRef]
127. Kimiz-Gebologlu I , Oncel SS . Exosomes: large-scale production, isolation, drug loading efficiency, and biodistribution and uptake. J Control Release. 2022; 347: 533– 43. doi:10.1016/j.jconrel.2022.05.027. [Google Scholar] [CrossRef]
128. Benjamin-Davalos S , Koroleva M , Allen CL , Ernstoff MS , La Shu S . Co-isolation of cytokines and exosomes: implications for immunomodulation studies. Front Immunol. 2021; 12: 638111. doi:10.3389/fimmu.2021.638111. [Google Scholar] [CrossRef]
129. Caruana JC , Dean SN , Walper SA . Isolation and characterization of membrane vesicles from Lactobacillus Species. Bio Protoc. 2021; 11( 17): e4145. doi:10.21769/BioProtoc.4145. [Google Scholar] [CrossRef]
130. Huang LH , Rau CS , Wu SC , Wu YC , Wu CJ , Tsai CW , et al. Identification and characterization of hADSC-derived exosome proteins from different isolation methods. J Cellular Molecular Medi. 2021; 25( 15): 7436– 50. doi:10.1111/jcmm.16775. [Google Scholar] [CrossRef]
131. He L , Zhu D , Wang J , Wu X . A highly efficient method for isolating urinary exosomes. Int J Mol Med. 2019; 43( 1): 83– 90. doi:10.3892/ijmm.2018.3944. [Google Scholar] [CrossRef]
132. Barathan M , Ng SL , Lokanathan Y , Ng MH , Law JX . The profound influence of gut microbiome and extracellular vesicles on animal health and disease. Int J Mol Sci. 2024; 25( 7): 4024. doi:10.3390/ijms25074024. [Google Scholar] [CrossRef]
133. Aberdam E , Closs B , Bordes S , Le Riche A , Aberdam D , Park BS . Extracellular vesicles including exosomes for hair follicle regeneration. In: Hair follicle regeneration. Berlin/Heidelberg, Germany: Springer; 2022. p. 205– 18. doi:10.1007/978-3-030-98331-4_9. [Google Scholar] [CrossRef]
134. Sharma A , Yadav A , Nandy A , Ghatak S . Insight into the functional dynamics and challenges of exosomes in pharmaceutical innovation and precision medicine. Pharmaceutics. 2024; 16( 6): 709. doi:10.3390/pharmaceutics16060709. [Google Scholar] [CrossRef]
135. Lee KWA , Chan LKW , Hung LC , Phoebe LKW , Park Y , Yi KH . Clinical applications of exosomes: a critical review. Int J Mol Sci. 2024; 25( 14): 7794. doi:10.3390/ijms25147794. [Google Scholar] [CrossRef]
136. Rezaie J , Feghhi M , Etemadi T . A review on exosomes application in clinical trials: perspective, questions, and challenges. Cell Commun Signal. 2022; 20( 1): 145. doi:10.1186/s12964-022-00959-4. [Google Scholar] [CrossRef]
137. Yadegar A , Bar-Yoseph H , Monaghan TM , Pakpour S , Severino A , Kuijper EJ , et al. Fecal microbiota transplantation: current challenges and future landscapes. Clin Microbiol Rev. 2024; 37( 2): e00060– 22. doi:10.1128/cmr.00060-22. [Google Scholar] [CrossRef]
138. Wang J , Liu X , Li Q . Interventional strategies for ischemic stroke based on the modulation of the gut microbiota. Front Neurosci. 2023; 17: 1158057. doi:10.3389/fnins.2023.1158057. [Google Scholar] [CrossRef]
139. Feng W , Liu J , Ao H , Yue S , Peng C . Targeting gut microbiota for precision medicine: focusing on the efficacy and toxicity of drugs. Theranostics. 2020; 10( 24): 11278– 301. doi:10.7150/thno.47289. [Google Scholar] [CrossRef]
140. Ryu G , Kim H , Koh A . Approaching precision medicine by tailoring the microbiota. Mamm Genome. 2021; 32( 4): 206– 22. doi:10.1007/s00335-021-09859-3. [Google Scholar] [CrossRef]
141. Shi B , Li H , He X . Advancing lifelong precision medicine for cardiovascular diseases through gut microbiota modulation. Gut Microbes. 2024; 16( 1): 2323237. doi:10.1080/19490976.2024.2323237. [Google Scholar] [CrossRef]
142. Guermazi D , Khan S , Shah A , Saliba E . Tiny messengers, big results: a review of exosome-mediated treatments and considerations in dermatology. Gene Protein Dis. 2024; 3( 2): 3230. doi:10.36922/gpd.3230. [Google Scholar] [CrossRef]
143. Kaltsas A , Zachariou A , Markou E , Dimitriadis F , Sofikitis N , Pournaras S . Microbial dysbiosis and male infertility: understanding the impact and exploring therapeutic interventions. J Pers Med. 2023; 13( 10): 1491. doi:10.3390/jpm13101491. [Google Scholar] [CrossRef]
144. Li Y , Wang YN , Luo YG , Yang H , Ren J , Li X . Advances in synthetic biology-based drug delivery systems for disease treatment. Chin Chem Lett. 2024; 35( 11): 109576. doi:10.1016/j.cclet.2024.109576. [Google Scholar] [CrossRef]
145. Green AA . Synthetic bionanotechnology: synthetic biology finds a toehold in nanotechnology. Emerg Top Life Sci. 2019; 3( 5): 507– 16. doi:10.1042/ETLS20190100. [Google Scholar] [CrossRef]
146. Mishra S , Tejesvi MV , Hekkala J , Turunen J , Kandikanti N , Kaisanlahti A , et al. Gut microbiome-derived bacterial extracellular vesicles in patients with solid tumours. J Adv Res. 2025; 68: 375– 86. doi:10.1016/j.jare.2024.03.003. [Google Scholar] [CrossRef]
147. Hersi F , Elgendy SM , Al Shamma SA , Altell RT , Sadiek O , Omar HA . Cancer immunotherapy resistance: the impact of microbiome-derived short-chain fatty acids and other emerging metabolites. Life Sci. 2022; 300: 120573. doi:10.1016/j.lfs.2022.120573. [Google Scholar] [CrossRef]
Cite This Article
 Copyright © 2026 The Author(s). Published by Tech Science Press.
Copyright © 2026 The Author(s). Published by Tech Science Press.This work is licensed under a Creative Commons Attribution 4.0 International License , which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
 Downloads
Downloads
 Citation Tools
Citation Tools